Под комплементарностью (взаимодополняемостью) понимают такое соответствие молекул биополимеров, которое обеспечивает возникновение между ними водородной связи.
В ДНК такое соответствие обеспечивается формированием парных связей (аденин-тимин и гуанин-цитозин).
Это необходимо для хранения и трансляции всей генетической информации человека, а заодно таких фундаментальных процессов, как репликации, транскрипции во время синтеза белка и возобновления этой кислоты вследствие повреждения ее цепи.
Описание комплементарности
Образование двойной цепи дезоксирибонуклеиновой кислоты (рнк) возможно тогда, когда одно пуриновое основание (аденин, гуанин) связано между собой одним из пиримидиновых оснований (тимином, цитозином.). Это соотношение известно как правило взаимодополняемости. Это правило означает, что две цепи несут одну и ту же генетическую информацию, хотя и различаются между собой химически. Получается, что одна цепь дезоксирибонуклеиновой кислоты задает другую.Дополняемость нуклеотидов обеспечивает важнейшую функцию нуклеинового соединения – определение синтеза белка. Вся информация о составе белка кодируется этими четырьмя основаниями – аденином, тимином, гуанином и цитозином. Образуется нуклеотидная последовательность, которая безошибочно передается от одного поколения к другому. По такому принципу происходит формирование идентичной молекулы – репликация. В свою очередь, нуклеотиды являются носителями всей информации, ведь каждая цепь служит своеобразной матрицей для получения новой.
История открытия РНК
Этот принцип был открыт Эрвином Чаргаффом в 1950 году. Но еще задолго до этого – в 1868 г.
было открыто РНК, а за шесть лет до открытия принципа взаимодополняемости было доказано, что именно эта кислота есть носителем генетической информации.
Чаргафф показал, что вследствие комплементарности нуклеотидов структуры молекул ДНК и РНК химически и геометрически соответствуют друг другу. Это было огромным прорывом в изучении наследственности, расшифровке дезоксирибонуклеиновой кислоты.
Принцип действия
Основы действия этого явления можно описать правилом Чаргаффа, которое гласит: Количество пуриновых основ (аденина и гуанина) равно содержанию пиримидиновых (тимина и цитозина);Количество аденина равняется содержанию тимина;Содержание гуанина равно соответственно цитозину. Немного позже А. Белозерский установил, что количественное соотношение пуриновых и пиримидиновых оснований есть постоянным для каждого отдельного вида организма. Иными словами, это соотношение является видовой характеристикой организма.
Для чего нужен принцип взаимодополняемости
Взаимодополняемость является важнейшей в формировании белков. Без нее невозможен синтез дочерней молекулы кислоты, которая была бы идентичной материнской. Без нее невозможно было представить себе деление клеток, ведь в ходе деления материнской клетки каждая новая клетка получает по одной копии ДНК, которая есть всегда одинаковой. Комплементарность обеспечивает передачу генетической информации от поколения к поколению. Это же позволяет понять механизм образования мутаций, а также способов их предотвращения. Изучения взаимодополняемости дало основание утверждать, что репликация дезоксирибонуклеиновой кислоты является важнейшим событием для деления клетки, синтеза белка. Использование комплементарности в генетике и медицинеЭто явление сегодня очень широко используется для внедрения в практическую медицину ДНК-технологий. Оно позволило подробнее изучить механизм развития наследственных болезней, анализировать основы их патогенеза. Вот некоторые области медицины и генетики, где успешно применяется такой принцип: Благодаря современным методам молекулярной медицины были созданы вакцины для борьбы с некоторыми формами гепатита, создан человеческий инсулин;Стало возможным восстанавливать нормальную свертываемость крови у больных гемофилией;В организм человека можно вводить полноценные гены, их фрагменты и таким образом корригировать некоторые нарушения обмена веществ;Стала возможной терапия некоторых форм иммунодефицита у детей;Разрабатываются эффективные методики лечения больных фенилкетонурией, муковисцидозом, гиперхолестеринемией и другими тяжелыми наследственными болезнями;Проводятся исследования человеческих генов.
 которая была бы идентичной материнской. Без нее невозможно было представить себе деление клеток, ведь в ходе деления материнской клетки каждая новая клетка получает по одной копии ДНК, которая есть всегда одинаковой. Комплементарность обеспечивает передачу генетической информации от поколения к поколению. Это же позволяет понять механизм образования мутаций, а также способов их предотвращения. Изучения взаимодополняемости дало основание утверждать, что репликация дезоксирибонуклеиновой кислоты является важнейшим событием для деления клетки, синтеза белка. Использование комплементарности в генетике и медицинеЭто явление сегодня очень широко используется для внедрения в практическую медицину ДНК-технологий. Оно позволило подробнее изучить механизм развития наследственных болезней, анализировать основы их патогенеза. Вот некоторые области медицины и генетики, где успешно применяется такой принцип: Благодаря современным методам молекулярной медицины были созданы вакцины для борьбы с некоторыми формами гепатита, создан человеческий инсулин;Стало возможным восстанавливать нормальную свертываемость крови у больных гемофилией;В организм человека можно вводить полноценные гены, их фрагменты и таким образом корригировать некоторые нарушения обмена веществ;Стала возможной терапия некоторых форм иммунодефицита у детей;Разрабатываются эффективные методики лечения больных фенилкетонурией, муковисцидозом, гиперхолестеринемией и другими тяжелыми наследственными болезнями;Проводятся исследования человеческих генов.
которая была бы идентичной материнской. Без нее невозможно было представить себе деление клеток, ведь в ходе деления материнской клетки каждая новая клетка получает по одной копии ДНК, которая есть всегда одинаковой. Комплементарность обеспечивает передачу генетической информации от поколения к поколению. Это же позволяет понять механизм образования мутаций, а также способов их предотвращения. Изучения взаимодополняемости дало основание утверждать, что репликация дезоксирибонуклеиновой кислоты является важнейшим событием для деления клетки, синтеза белка. Использование комплементарности в генетике и медицинеЭто явление сегодня очень широко используется для внедрения в практическую медицину ДНК-технологий. Оно позволило подробнее изучить механизм развития наследственных болезней, анализировать основы их патогенеза. Вот некоторые области медицины и генетики, где успешно применяется такой принцип: Благодаря современным методам молекулярной медицины были созданы вакцины для борьбы с некоторыми формами гепатита, создан человеческий инсулин;Стало возможным восстанавливать нормальную свертываемость крови у больных гемофилией;В организм человека можно вводить полноценные гены, их фрагменты и таким образом корригировать некоторые нарушения обмена веществ;Стала возможной терапия некоторых форм иммунодефицита у детей;Разрабатываются эффективные методики лечения больных фенилкетонурией, муковисцидозом, гиперхолестеринемией и другими тяжелыми наследственными болезнями;Проводятся исследования человеческих генов.Перспективы развития исследований
На современном этапе развития медицины и генетики взаимодополняемость получает свое широкое применение во многих исследованиях. Так, она позволяет устанавливать, внедрять в лечебную практику такие основополагающие принципы функционирования живых организмов, как саморегуляция, взаимоотношение функциональных систем, организации функций и проч.
Комплементарность позволяет использовать такие методы лечения, которые были бы направлены как бы «внутрь» организма, с использованием его компенсаторных возможностей.
Изучение нуклеотидов дает большие возможности внедрять в основные методы лечения самые последние достижения генной инженерии с тем, чтобы побороть тяжелые наследственные болезни и обеспечить больным полноценную жизнь.
Интересные факты о взаимодополняемости
В ходе исследований выяснились такие интересные факты: В человеческом геноме свыше трех миллиардов «букв» – нуклеотидов;Только лишь один их процент кодирует белки;Всего у человека свыше двадцати тысяч генов;Человеческий геном хранится в каждой(!) клетке;Около четырех пятых всего генома «переписывается» на РНК – рибонуклеиновую кислоту;В ДНК имеется огромное количество вспомогательных участков, контролирующих весь сложнейший процесс кодировки и синтеза белка.Впрочем, возможности комплементарности для изучения нашего генома до конца не исследованы, так что перед нами – новые открытия, связанные с генетикой.
 В человеческом геноме свыше трех миллиардов «букв» – нуклеотидов;Только лишь один их процент кодирует белки;Всего у человека свыше двадцати тысяч генов;Человеческий геном хранится в каждой(!) клетке;Около четырех пятых всего генома «переписывается» на РНК – рибонуклеиновую кислоту;В ДНК имеется огромное количество вспомогательных участков, контролирующих весь сложнейший процесс кодировки и синтеза белка.Впрочем, возможности комплементарности для изучения нашего генома до конца не исследованы, так что перед нами – новые открытия, связанные с генетикой.
В человеческом геноме свыше трех миллиардов «букв» – нуклеотидов;Только лишь один их процент кодирует белки;Всего у человека свыше двадцати тысяч генов;Человеческий геном хранится в каждой(!) клетке;Около четырех пятых всего генома «переписывается» на РНК – рибонуклеиновую кислоту;В ДНК имеется огромное количество вспомогательных участков, контролирующих весь сложнейший процесс кодировки и синтеза белка.Впрочем, возможности комплементарности для изучения нашего генома до конца не исследованы, так что перед нами – новые открытия, связанные с генетикой.Днк (дезоксирибонуклеиновая кислота)
ДНК (дезоксирибонуклеиновая кислота) — это линейный органический полимер, мономерными звеньями которого являются нуклиатиды.
Вся информация о строении и функционировании любого живого организма содержится в закодированном виде в его генетическом материале. Основу генетического материала организма составляет дезоксирибонуклеиновая кислота (ДНК).
ДНК большинства организмов – это длинная двухцепочечная полимерная молекула.
Последовательность мономерных звеньев (дезоксирибонуклеотидов) в одной ее цепи соответствует (комплементарна) последовательности дезоксирибонуклеотидов в другой.
Принцип комплементарности обеспечивает синтез новых молекул ДНК, идентичных исходным, при их удвоении (репликации).
Участок молекулы ДНК, кодирующий определенный признак, – ген.
Гены – это индивидуальные генетические элементы, имеющие строго специфичную нуклеотидную последовательность, и кодирующие определенные признаки организма. Одни из них кодируют белки, другие — только молекулы РНК.
Информация, которая содержится в генах, кодирующих белки (структурных генах), расшифровывается в ходе двух последовательных процессов:
- синтеза РНК (транскрипции): на определенном участке ДНК как на матрице синтезируется матричная РНК (мРНК).
- синтеза белка (трансляции): В ходе согласованной работы многокомпонентной системы при участии транспортных РНК (тРНК), мРНК, ферментов и различных белковых факторов осуществляется синтез белковой молекулы.
Все эти процессы обеспечивают правильный перевод зашифрованной в ДНК генетической информации с языка нуклеотидов на язык аминокислот. Аминокислотная последовательность белковой молекулы определяет ее структуру и функции.
Строение ДНК
ДНК – это линейный органический полимер. Его мономерные звенья – нуклеотиды, которые, в свою очередь, состоят из:
- азотистого основания;
- пятиуглеродного сахара (пентозы);
- фосфатной группы (рисунок 1).
Рисунок 1 : ДНК – строение одной цепочки нуклеотидов
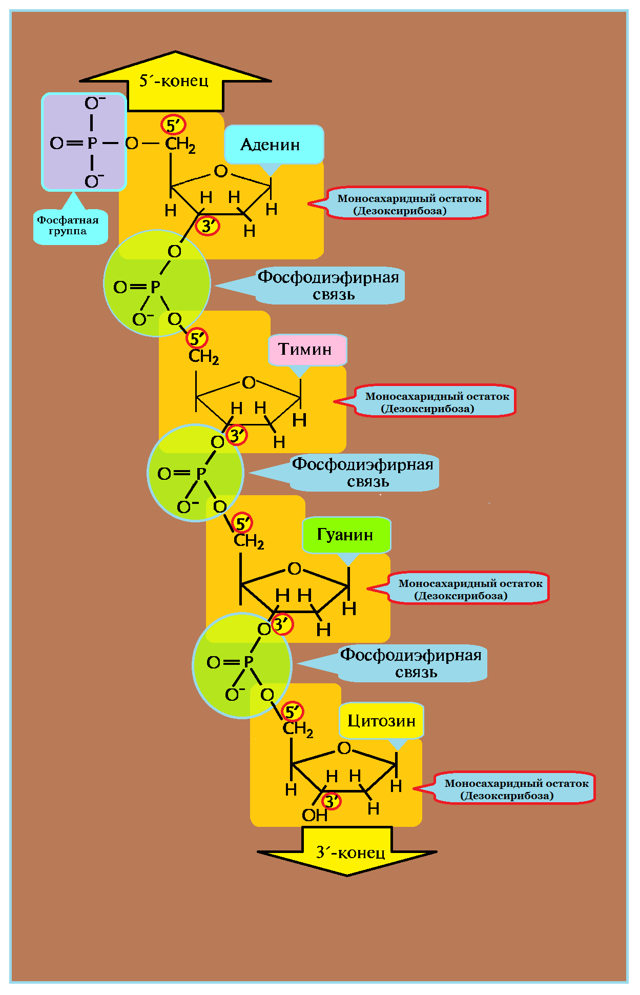 Рисунок 1 : ДНК – строение одной цепочки нуклеотидов
Рисунок 1 : ДНК – строение одной цепочки нуклеотидовПри этом, фосфатная группа присоединена к 5′-атому углерода моносахаридного остатка, а органическое основание — к 1′-атому.
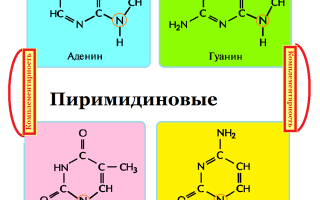
Основания в ДНК бывают двух типов:
- Пуриновые: аденин ( А ) и гуанин (G);
- Пиримидиновые: цитозин (С) и тимин (Т);(рисунок 2),
Рисунок 2: Азотистые основания- пуриновые и пиримидиновые
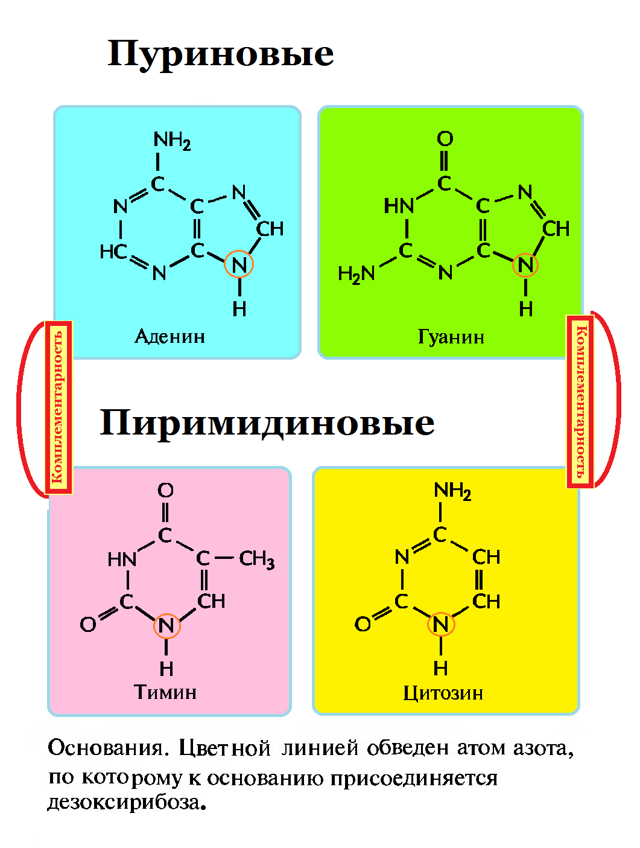 Рисунок 2: Азотистые основания- пуриновые и пиримидиновые
Рисунок 2: Азотистые основания- пуриновые и пиримидиновыеСтроение нуклеотидов в молекуле ДНК
В ДНК моносахарид представлен 2′-дезоксирибозой, содержащей только 1 гидроксильную группу (ОН), а в РНК — рибозой, имеющей 2 гидроксильные группы (OH).
Нуклеотиды соединены друг с другом фосфодиэфирными связями, при этом фосфатная группа 5′-углеродного атома одного нуклеотида связана с З’-ОН-группой дезоксирибозы соседнего нуклеотида (рисунок 1). На одном конце полинуклеотидной цепи находится З’-ОН-группа (З’-конец), а на другом — 5′-фосфатная группа (5′-конец).
Уровни структуры ДНК
Принято выделять 3 уровня структуры ДНК:
- первичную;
- вторичную;
- третичную.
- Первичная структура ДНК – это последовательность расположения нуклеотидов в полинуклеотидной цепи ДНК.
- Вторичная структура ДНК стабилизируется водородными связями между комплементарными парами оснований и представляет собой двойную спираль из двух антипараллелных цепочек, закрученных вправо вокруг одной оси.
- Общий виток спирали- 3,4нм, расстояние между цепочками 2нм.
Третичная структура ДНК – суперсперализация ДНК.
Двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации с образованием суперспирали или открытой кольцевой формы, что часто вызвано ковалентным соединением их открытых концов.
Суперспиральная структура ДНК обеспечивает экономную упаковку очень длинной молекулы ДНК в хромосоме. Так, в вытянутой форме длина молекулы ДНК составляет 8 см, а в форме суперспирали укладывается в 5 нм.
Правило Чаргаффа
Правило Э. Чаргаффа – это закономерность количественного содержания азотистых оснований в молекуле ДНК:
- У ДНК молярные доли пуриновых и пиримидиновых оснований равны: А+ G = C + Т или (А + G)/(C + Т)=1.
- В ДНК количество оснований с аминогруппами (А +C) равно количеству оснований с кетогруппами (G + Т): А +C= G + Т или (А +C)/(G + Т)= 1
- Правило эквивалентности, то есть : А=Т, Г=Ц; А/Т = 1; Г/Ц=1.
- Нуклеотидный состав ДНК у организмов различных групп специфичен и характеризуется коэффициентом специфичности: (Г+Ц)/(А+Т). У высших растений и животных коэффициент специфичности меньше 1, и колеблется незначительно: от 0,54 до 0,98, у микроорганизмов он больше 1.
Модель ДНК Уотсона-Крика
Б 1953 г. Джеймс Уотсон и Фрэнсис Крик, основываясь на данных рентгеноструктурного анализа кристаллов ДНК, пришли к выводу, что нативная ДНК состоит из двух полимерных цепей, образующих двойную спираль (рисунок 3).
Навитые одна на другую полинуклеотидные цепи удерживаются вместе водородными связями, образующимися между комплементарными основаниями противоположных цепей (рисунок 3).
При этом аденин образует пару только с тимином, а гуанин — с цитозином.
Пара оснований А—Т стабилизируется двумя водородными связями, а пара G—С — тремя.
Длина двухцепочечной ДНК обычно измеряется числом пар комплементарных нуклеотидов (п.н.). Для молекул ДНК, состоящих из тысяч или миллионов пар нуклеотидов, приняты единицы т.п.н. и м.п.н. соответственно. Например, ДНК хромосомы 1 человека представляет собой одну двойную спираль длиной 263 м.п.н.
Сахарофосфатный остов молекулы, который состоит из фосфатных групп и дезоксирибозных остатков, соединенных 5’—З’-фосфодиэфирными связями, образует «боковины винтовой лестницы», а пары оснований А—Т и G—С — ее ступеньки (рисунок 3).
Рисунок 3: Модель ДНК Уотсона-Крика
Цепи молекулы ДНК антипараллельны: одна из них имеет направление 3’→5′, другая 5’→3′.
В соответствии с принципом комплементарности, если в одной из цепей имеется нуклеотидная последовательность 5-TAGGCAT-3′, то в комплементарной цепи в этом месте должна находиться последовательность 3′-ATCCGTA-5′. В этом случае двухцепочечная форма будет выглядеть следующим образом:
- 5′-TAGGCAT-3′
- 3-ATCCGTA-5′.
- В такой записи 5′-конец верхней цепи всегда располагают слева, а 3′-конец — справа.
- Носитель генетической информации должен удовлетворять двум основным требованиям: воспроизводиться (реплицироваться) с высокой точностью и детерминировать (кодировать) синтез белковых молекул.
- Модель ДНК Уотсона—Крика полностью отвечает этим требованиям, так как:
- согласно принципу комплементарности каждая цепь ДНК может служить матрицей для образования новой комплементарной цепи. Следовательно, после одного раунда репликации образуются две дочерние молекулы, каждая из которых имеет такую же нуклеотидную последовательность, как исходная молекула ДНК.
- нуклеотидная последовательность структурного гена однозначно задает аминокислотную последовательность кодируемого ею белка.
Интересные факты о ДНК
- Одна молекула ДНК человека вмещает порядка 1,5 гигабайта информации. При этом, ДНК всех клеток человеческого организма занимают 60 млрд. терабайт, что сохраняются на 150-160 граммах ДНК. [2]
- Международный день ДНК отмечается 25 апреля.
Именно в этот день в 1953 году Джеймс Уотсон и Фрэнсис Крик опубликовали в журнале Nature свою статью под названием «Молекулярная структура нуклеиновых кислот», где описали двойную спираль молекулы ДНК.
[3]
Список литературы: Молекулярная биотехнология: принципы и применение, Б.Глик, Дж. Пастернак, 2002 год Б.Глик, Дж. Пастернак, Источник: Молекулярная биотехнология: принципы и применение, Б.Глик, Дж. Пастернак, 2002 год [2] MPlast.
by – портал: “ДНК 1 клетки человека вмещает 1,5 гигабайта информации – лучший винчестер на планете” – 27 апреля 2016 года [3] Журнал NATURE: “Molecular Structure of Nucleic Acids” – 25 апреля 1953 года Дата в источнике: 2002 год
Дезоксирибонуклеиновые кислоты (ДНК) и Принцип комплементарности • биология-в.рф
Дезоксирибонуклеиновые кислоты (ДНК) и Принцип комплементарности

Дезоксирибонуклеиновые кислоты (ДНК)
Основная часть ДНК сосредоточена в хромосомах клетки. Незначительная ее часть находится в митохондриях и пластидах. У прокариот – кольцевая ДНК, плазмиды. ДНК – это длинная полинуклеотидная цепь, которая может достигать 10 м. Соответственно, она имеет большую молекулярную массу. В ядре ДНК размещается благодаря образованию компактной структуры.
Происходит это из-за взаимодействия с основными белками – гистонами (от греч. гистос – ткань). Гистоны одинаковые почти у всех эукариот. Двойная спираль ДНК намотана на молекулы пистонов, образующих сердцевину, и сокращается почти в пять раз. Образуется нуклеосома (от лат .nucleus – ядро от греч. сома – тело).
Нуклеосомы взаимодействуют с другими белками и образуют спираль высшего уровня.
Первооткрыватели структуры ДНК
Американский биохимик Дж. Уотсон и английский генетик Ф. Крик в 1953 году предложили модель пространственной структуры ДНК. Позднее она была подтверждена экспериментально. ДНК состоит из двух полинуклеотидных цепей.
Они соединены между собой водородными связями, которые образуются между нуклеотидами: два – между аденином и тимином, три – между цитозином и гуанином. Каждый нуклеотид расположен в плоскости, которая перпендикулярна оси спирали.
Две полинуклеотидные цепи ДНК, в соответствии с предложенной моделью, образуют правозакрученую спираль – вторичную структуру ДНК. Спирали ДНК разнонаправлены. Между азотистыми основаниями расстояние составляет 0,34 нм.
Шаг спирали равняется 3,4 нм и содержит десять пар азотистых оснований, диаметр спирали составляет около 2 нм. Спираль ДНК может испытывать пространственное уплотнение и формировать суперспираль – третичную структуру.
Это возможно потому, что ДНК соединяется с ядерными белками.
Последним уровнем структурной организации ДНК является хромосома, которая наиболее компактна в период деления. У многих прокариот, некоторых вирусов, в митохондриях и хлоропластах эукариот ДНК имеет кольцевую структуру и не соединяется с белками.
Американский ученый Э. Чаргафф и его коллеги в 1950 году исследовали состав ДНК и обнаружили определенные закономерности. Правило Э. Чаргаффа (для ДНК): число адениловых нуклеотидов равняется количеству тимидиловых, а гуаниловых количеству цитозидиловых, то есть А = Т, Г = Ц, или А + Г (Ц) = Т + Ц (Г). Соотношение (А + Г)/ (Т + Ц) = 1.
Принцип комплементарности
Принцип комплементарности (от лат. complementum – дополнение) – это правило строгого соответствия: аденина (А) – тимину (Т) или урацилу (У), гуанина (Г) – цитозину (Ц).
Молекулы ДНК способны к самоудвоению – репликации (от англ. replica – отражение), редупликации (от лат. duplicatio– удвоение). В основе ее лежит принцип комплементарности. В новообразованной цепи последовательность нуклеотидов определяется их последовательностью в цепи первичной молекулы ДНК.
Одна цепь принадлежит материнской молекуле, вторая – синтезируется. Такой метод называется полуконсервативным. Синтез новых цепей катализируется ферментом ДНК-полимеразой. Фермент этот движется вдоль каждой цепи от З’-конца до 5′-конца. Репликация начинается одновременно на многих участках ДНК.
Синтезированные участки ДНК собираются потом в единое целое. Удвоение происходит в синтетический период интерфазы. Скорость репликации ДНК у млекопитающих составляет приблизительно 50 нуклеотидов в секунду. Под влиянием внешних факторов могут возникнуть любые повреждения. Они могут быть исправлены на матрице второй цепи.
Соответствующие ферменты удаляют поврежденные участки, встраивают новые. Этот процесс называется репарацией.
Двухспиральная ДНК полностью или частично может распадаться на отдельные цепи и терять биологическую активность при определенных условиях (действие кислот, щелочей, высоких или низких температур и т. п.).
Происходит процесс денатурации ДНК, то есть разрыв водородных связей между комплементарными азотистыми основаниями.
После прекращения действия факторов ДНК может восстановить двухспиральное строение, то есть может происходить процесс ренатурации.
Функции ДНК
Сохраняет, воссоздает наследственную информацию, во время деления материнской клетки обеспечивает ее передачу дочерним клеткам, из нее происходит синтез белка. Количество ДНК в клетке является видовым признаком.
Молекулярный уровеньУровни организации живого
ЕГЭ. Принцип комлементарности. Репликация ДНК
Приветствую друзья, продолжаем разбирать теорию связанную с генетическим кодом клетки. В прошлой статье мы узнали как клетка хранит и использует свой генетический материал, а сегодня поговорим о процессах, которые сопутствуют этому.
Мы помним, что гены – это всего лишь информация о структуре каких-либо белков. Каждый ген (небольшой участок ДНК) отвечает за их сборку (синтез) на рибосомах (органеллы, где происходит сборка белков из аминокислот).
Процессы транскрипции и трансляции в клетке (с) картинка из интернета Процессы транскрипции и трансляции в клетке (с) картинка из интернета
Все эти процессы: удвоения ДНК (репликация), переписывания ДНК на информационную РНК (транскрипция) и сборки белка из аминокислот в рибосомах за счёт тРНК (трансляция) и называются реакциями матричного синтеза.
Реакции матричного синтеза. Репликация. Транскрипция. Трансляция. Реакции матричного синтеза. Репликация. Транскрипция. Трансляция.
Не стоит бояться в этой схеме словосочетания “нуклеиновые кислоты” – это просто обобщённое название для всех ДНК и РНК в клетке. Они так называются, потому что чаще всего находятся в ядре (от лат. nucleus — ядро).
Принцип комплементарности и репликация ДНК
Поскольку процесс создания (синтеза) белка начинается с процесса переписывания (транскрипция) его структуры молекулой иРНК с ДНК, то очень важно, чтобы запись начиналась и заканчивалась в строго определенных местах. Тогда получившийся белок будет “полноценным” и сможет корректно выполнять свои биологические функции.
Для этого и существует принцип комплементарности. Он позволяет парам нуклеотидов (из которых и состоит ДНК) связываться только с подходящими для них молекулами.
Принцип комплементарности для ДНК. Тимин комплементарен аденину (двойная связь), гуанин комплементарен цитозину (тройная связь) (с) картинка из интернета Принцип комплементарности для ДНК. Тимин комплементарен аденину (двойная связь), гуанин комплементарен цитозину (тройная связь) (с) картинка из интернета
У РНК есть свои особенности, это необходимо для того, чтобы клетка могла отличить её от ДНК, поэтому в ней молекулы Тимина (Т) заменены на молекулы Урацила (У). Получается, что при переписывании (транскрипции) гена вместо пары А=Т, получается пара А=У, остальное происходит без изменений. Посмотрите пример ниже для лучшего понимания.
Принцип комплементарности для РНК. Аденин комплиментарен Урацилу. Гуанин комлементарен Цитозину. Принцип комплементарности для РНК. Аденин комплиментарен Урацилу. Гуанин комлементарен Цитозину.
При делении клетки, ей необходимо удвоить свою генетическую информацию (для того, чтобы каждая новая клетка имела полный набор генов).
Это и называется процессом репликации ДНК и возможно как раз таки за счёт принципа комлементарности.
Двойную цепь ДНК можно развернуть и достроить парными нуклеотидами получив уже две двойных (сестринских) цепи, которые будут абсолютно идентичны.
Процесс репликации. Удвоении ДНК клеткой перед делением (с) картинка из интернета Процесс репликации. Удвоении ДНК клеткой перед делением (с) картинка из интернета
Очень важно разбираться в процессах матричного синтеза. Данная тема обширно представлена в ЕГЭ как в B части, где требуется указать процентное соотношение тех или иных нуклеотидов в ДНК/РНК, так и в C части, где просят достроить комплементарную цепь и посмотреть какие белки на РНК в ней закодированы.
Эти и другие задания мы обязательно разберём на канале, а пока подписывайтесь, ставьте лайки (это продвигает публикацию и образовывает больше людей) и делитесь своими вопросами и мнением в х. Всем удачи 🙂
ДЕЗОКСИРИБОНУКЛЕИ́НОВЫЕ КИСЛО́ТЫ
Авторы: А. А. Богданов
ДЕЗОКСИРИБОНУКЛЕИ́НОВЫЕ КИСЛО́ТЫ (ДНК), нуклеиновые кислоты, содержащие в качестве углеводного компонента дезоксирибозу. ДНК – осн.
компонент хромосом всех живых организмов, вещество, из которого построены геномы всех про- и эукариот, а также внехромосомные наследств. элементы (плазмиды) и геномы мн. вирусов.
В клетках прокариот ДНК организована в виде компактного образования – нуклеоида. У эукариот она содержится в ядрах клеток и в органеллах – митохондриях и хлоропластах. В нуклеотидной последовательности ДНК записана (кодирована) генетич.
информация о всех признаках вида и особенностях индивидуума. Все осн. генетич. процессы – репликация, транскрипция и рекомбинация связаны с функционированием молекулы ДНК.
Впервые ДНК в виде комплексов с белками (дезоксирибонуклеопротеидов) была открыта в 1868 И. Ф. Мишером в ядрах клеток гноя и сперме рыб. Долгое время считалось, что ДНК содержится только в клетках животных, и лишь к сер. 1930-х гг. было доказано (А. Н.
Белозерский), что ДНК – непременный компонент каждой живой клетки. В 1944 амер. микробиолог О. Эйвери с сотрудниками показали, что с помощью ДНК, тщательно очищенной от всех остальных клеточных компонентов, наследуемый биологич. признак может быть перенесён из одной клетки в другую.
Тем самым была определена биологич. функция ДНК как вещества наследственности.
В кон. 19 – нач. 20 вв.
было установлено, что ДНК представляют собой полимерные молекулы, мономерными составляющими которых служат дезоксирибонуклеотиды, содержащие остатки дезоксирибозы, фосфорной кислоты и одно из четырёх азотистых оснований: пуриновых – гуанина (G) и аденина (А) и пиримидиновых – цитозина (C) и тимина (Т). В кон. 1940-х – нач. 1950-х гг. в лаборатории А. Тодда было доказано, что единственным типом межнуклеотидной связи в полимерных цепях ДНК является 3'–5'-фосфодиэфирная связь. В это же время Э. Чаргафф с сотрудниками выяснили осн. закономерности нуклеотидного состава ДНК (правила Чаргаффа), наиболее важная из которых – равенство содержания остатков аденина и тимина (А = Т), а также гуанина и цитозина (G = C).
Основываясь на этих данных, в 1953 Дж. Уотсон и Ф. Крик расшифровали рентгенограммы кристаллов ДНК, полученные в лабораториях Р. Франклин и М. Уилкинса, и сделали одно из выдающихся открытий совр. естествознания.
Они установили, что молекула ДНК представляет собой регулярную спираль, состоящую из двух полинуклеотидных цепей (двойная спираль). Диаметр спирали постоянен на протяжении всей её длины и равен примерно 2 нм. Длина витка спирали составляет 3,4 нм. На один виток в одной цепи приходится примерно 10 нуклеотидных остатков, т. е.
межнуклеотидное расстояние вдоль оси спирали равно 0,34 нм. Азотистые основания в двойной спирали ДНК лежат в одной плоскости, которая практически перпендикулярна её гл. оси.
При этом основания, принадлежащие разным цепям и находящиеся напротив друг друга, образуют комплементарные пары, стабилизированные водородными связями таким образом, что аденин всегда спарен только с тимином, а гуанин – с цитозином (пары G – C связаны между собой тремя водородными связями, а пары А – Т лишь двумя).
Для стабилизации структуры двойной спирали ДНК важное значение имеют также взаимодействия между плоскостями соседних оснований, принадлежащих одной и той же цепи (т. н. стэкинг-взаимодействия, от англ. stack – стог, складывать в стог, располагать один над другим).
Из модели двойной спирали Уотсона – Крика прямо вытекает принцип самовоспроизведения (удвоения, репликации) молекулы ДНК (а следовательно, и любого генетич.
материала): если две комплементарные цепи ДНК разделить, а затем на каждой, как на матрице, построить новые, строго комплементарные им цепи, то две дочерние двуспиральные молекулы будут идентичны материнской.
Открытие этого принципа позволило на молекулярном уровне объяснить явление наследственности и положило начало молекулярной биологии. Принцип комплементарного спаривания оснований нуклеиновых кислот лежит в основе всех процессов передачи генетич. информации в клетке.
В двойной спирали ДНК сахарофосфатный остов полинуклеотидных цепей обращён наружу, а на поверхности спирали можно выделить две бороздки: большую – шириной 2,2 нм и малую – шириной 1,2 нм. Двойная спираль ДНК, описанная Дж.
Уотсоном и Ф. Криком, – правозакрученная, а полинуклеотидные цепи в ней антипараллельны, т. е. направлены в противоположные стороны, так что 3'-конец одной цепи располагается напротив 5'-конца другой.
Она была названа В-формой ДНК.
Оказалось, однако, что двойная спираль ДНК характеризуется существенным полиморфизмом и при изменении внешних условий может принимать пространственную структуру (конформацию), отличную от уотсон-криковской В-формы. Так, при понижении влажности в препарате или, напр.
, при добавлении спирта к водному раствору ДНК она переходит в т. н.
А-форму, отличающуюся от В-формы шириной и глубиной бороздок, увеличением диаметра спирали, смещением пар оснований к периферии спирали и их заметным наклоном по отношению к оси спирали, а внутри неё образуется полость диаметром 0,4 нм.
В основе этих структурных превращений лежит изменение конформации остатка дезоксирибозы, что, в свою очередь, ведёт к изменению расстояния между фосфатными группами соседних нуклеотидных остатков одной цепи.
При высокой концентрации солей участки двойных спиралей ДНК с чередующимися нуклеотидными последовательностями типа многократно повторяющегося гуанозин-цитозинового динуклеотида (GC) из правозакрученной формы переходят в левозакрученную.
У этой формы ДНК линия, соединяющая фосфатные группы, через каждые две пары имеет излом и принимает зигзагообразный вид. Такая конформация ДНК называется Z-формой (от англ. zigzag). Хотя полиморфизм ДНК может играть существенную роль в регуляции активности генов, прямых данных о наличии у двойной спирали ДНК in vivo иных конформаций, кроме В-формы, пока нет.
Важным свойством двойных спиралей ДНК является их микрогетерогенность, обнаруживаемая рентгеноструктурным анализом высокого разрешения.
Она обусловлена тонкими различиями в конформации нуклеотидных остатков, появление которых зависит от последовательности расположения нуклеотидов в цепи, и проявляется в образовании характерных изгибов и изломов.
Такие особенности структуры молекулы ДНК, несомненно, должны быть связаны с её функционированием.
При наличии в молекуле ДНК повторяющихся последовательностей (палиндромов) могут формироваться пары не только между основаниями противоположных цепей, но и в пределах одной цепи, что создаёт возможность образования связанных водородными связями своеобразных шпилек с петлями.
При повышении темп-ры или рН растворов ДНК, в присутствии ряда органич. веществ и др. соединений происходит денатурация ДНК – разрыв водородных связей между парами оснований и разрушение регулярной структуры двойной спирали, которое завершается полным разделением цепей.
Благодаря кооперативному характеру внутримолекулярных взаимодействий, стабилизирующих двойную спираль, этот процесс напоминает фазовый переход и поэтому называется плавлением ДНК. В условиях, оптимальных для образования двойной спирали, отд.
комплементарные цепи ДНК способны реассоциировать с восстановлением исходной двуспиральной структуры (ренатурация ДНК).
Это свойство лежит в основе метода молекулярной гибридизации нуклеиновых кислот, который позволяет выявлять степень сходства нуклеотидных последовательностей молекул ДНК или ДНК и РНК, особенности их организации, в т. ч. наличие и число повторов (см. Нуклеотидные последовательности).
Последовательность чередования нуклеотидных остатков в ДНК (первичная структура) у разных организмов строго индивидуальна и служит важнейшей характеристикой, отличающей одну молекулу ДНК от другой и соответственно один ген или один регуляторный генетич. элемент от другого.
Размеры молекул ДНК варьируют от нескольких тысяч пар нуклеотидов (т. п. н.) у плазмид и некоторых вирусов до сотен т. п. н. у высших организмов. Содержание ДНК в разных организмах также различно и по числу образующих её нуклеотидов составляет от 5·106 у бактерий до 2·1011 пар нуклеотидов (п. н.
) у высших растений (в расчёте на гаплоидный геном). Эти гигантские молекулы чрезвычайно компактно упакованы в клетках или вирусах. В прокариотич. нуклеотиде такая укладка поддерживается небольшим количеством спец. белков и, вероятно, рибонуклеиновыми кислотами (РНК). Описано неск.
уровней упаковки эукариотической ДНК с помощью универсального набора гистонов и некоторых негистоновых белков, приводящих к образованию осн. компонента хромосомы – хроматина. Напр.
, длина ДНК самой большой хромосомы человека равна 8 см, но в хромосоме (в состоянии митоза) она не превышает 5 мкм.
В ядрах эукариот (за исключением гамет) ДНК представлена двумя копиями. Каждая про- и эукариотическая хромосома содержит только одну молекулу двуспиральной ДНК.
Геном подавляющего большинства вирусов также представлен двуспиральной ДНК, и лишь некоторые фаги в качестве геномной содержат однотяжевую кольцевую или линейную молекулу ДНК.
В кольцо замкнуты молекулы двунитевых ДНК прокариотич. хромосомы, плазмид и мн. вирусов, ДНК митохондрий и хлоропластов. При этом если цепь ковалентно-непрерывна (т. е. все фосфодиэфирные связи замкнуты), то циклич.
ДНК могут находиться в сверхспирализованной форме, когда нити двойной спирали многократно зацеплены друг с другом. В клетке сверхвитки создаются и разрушаются ферментами топоизомеразами.
Циклическая сверхспирализованная ДНК обладает определённым запасом энергии по сравнению с её линейной формой, поэтому образование сверхвитков важно для функционирования ДНК (напр., позволяет разрешать топологич. трудности, возникающие при репликации).
Кроме того, благодаря наличию сверхвитков могут образовываться необычные структуры в её макромолекуле: крестообразные структуры (в палиндромах), Z-форма, тринитевые участки, или т. н. Н-форма (в гомопурин-гомопиримидиновых блоках).
Биосинтез ДНК (репликация) осуществляется путём матричного синтеза при участии ферментов ДНК-полимераз совместно с большой группой вспомогат. белков и находится под контролем спец. регуляторных систем клетки.
In vitro любой участок ДНК может быть амплифицирован с помощью полимеразной цепной реакции.
В ходе репликации in vivo, а также после её окончания происходит метилирование небольшого числа определённых остатков цитозина с образованием 5-метилцитозина, представляющее собой специфич.
процесс модификации ДНК, непосредственно связанный с её последующим функционированием. Метилирование и деметилирование ДНК играют важную роль в процессах эмбрио- и гаметогенеза.
В ходе жизнедеятельности организмов их ДНК под влиянием внешних факторов может подвергаться разл. повреждающим воздействиям, сопровождающимся нарушением структуры азотистых оснований. В ходе эволюции клетки выработали защитные механизмы, обеспечивающие восстановление исходной структуры – репарацию ДНК.
В клетке ДНК расщепляется специфич. ферментами – дезоксирибонуклеазами. Среди них наиболее известны эндонуклеазы рестрикции, защищающие клетку от чужеродной ДНК и широко применяемые в генетич. инженерии.
В нач. 1970-х гг. Ф. Сенгером и др. были разработаны эффективные методы определения последовательности нуклеотидов в молекулах ДНК (см. Секвенирование). В кон. 20 в. на основе этих методов создана мощная автоматизир.
технология секвенирования ДНК, с помощью которой определена первичная структура ДНК полных геномов мн. вирусов, митохондрий, хлоропластов, бактерий, растений и животных. К 2004 было завершено определение нуклеотидной последовательности практически всего генома человека (более трёх млрд. п. н.).
Эти работы стимулировали развитие биоинформатики и положили начало новому разделу молекулярной генетики – геномике.
Информация о нуклеотидных последовательностях ДНК широко используется при создании рекомбинантных ДНК – молекул с заданными свойствами, включающих структурные элементы ДНК разных организмов (см. Генетическая инженерия), а также при конструировании новых белков (см.
Белковая инженерия). Знание первичной структуры ДНК важно при анализе наследств. и онкологич. заболеваний, идентификации личности (см. ДНК-типирование), при амплификации и выделении определённых генов, регуляторных элементов и др. функционально важных участков ДНК.