Эукариотические клетки отличаются от бактериальных во многих отношениях. Они крупнее, содержат ядро, отделяющее геном и его транскрипцию от процессов цитоплазмы. В цитоплазме эукариот находятся сложные системы мембранных цистерн и пузырьков — эндоплазматический ретикулум, аппарат Гольджи, лизосомы и другие вакуоли.
Пузырьки управляемо отшнуровываются от мембран в одних местах и сливаются с ними в других. Белки цитоскелета — актин и тубулин — поддерживают форму клетки, а связанные с ними моторные белки — миозин, динеин, кинезин — перемещают органеллы внутри клетки и обеспечивают движение всей клетки.
Рибосомы эукариотической клетки крупнее, чем бактериальные, содержат больше белков, и для узнавания мРНК нуждаются в кэпе — специальной модификации 5' конца мРНК. Кроме кэпа, эукариотические мРНК имеют полиадениловый хвост на 3' конце, и кодируют только одну рамку считывания.
Дыхательные цепи и мембранные АТФазы убраны с плазматической мембраны в специальные органеллы — митохондрии и пластиды. Геном эукариот состоит из нескольких линейных молекул ДНК — хромосом, намотанных для компактности на специальные белковые «катушки» – гистоны. Размер генома может достигать десятков миллиардов пар нуклеотидов, что на четыре порядка больше, чем у бактерий.
Геном часто переполнен некодирующими последовательностями разных типов, из которых шире всего распространены сплайсеосомные интроны. При делении клетки цитоскелет разделяет реплицированные хромосомы по дочерним клеткам в сложном движении, напоминающем эпизод классического балета.
Наконец, в жизненном цикле многих эукариот есть половой процесс, в котором в одной клетке собираются гены двух родителей, и мейоз — специальное деление, в котором хромосомы, происходящие от разных родителей, обмениваются отдельными участками, после чего диплоидная клетка делится на четыре гаплоидных (с одиночным хромосомным набором).
Первым шагом в понимании происхождения эукариотической клетки стало обнаружение сходства между митохондриями и свободноживущими аэробными бактериями, а так же между пластидами и цианобактериями. Первые работы в этом направлении появились еще в конце XIX века, а в современном виде теорию симбиогенеза сформулировала Маргулис в 1970-ых годах.
К тому времени уже было обнаружено, что пластиды и митохондрии имеют собственную миниатюрную генетическую систему, включающую в себя кольцевую молекулу ДНК, особые рибосомы — мельче, чем в цитоплазме эукариот, и похожие на бактериальную — и никогда не образуются «с нуля», всегда происходя путем роста и деления существующих. Все это указывает на происхождение пластид и митохондрий от бактериальных симбионтов, когда-то поселившихся в цитоплазме эукариотической клетки.
Тогда же, в 1970-ые, были попытки расширить теорию симбиогенеза и на другие системы эукариотической клетки. Несколько раз сообщалось об обнаружении собственной ДНК в центриолях — центрах огранизации тубулиновго цитоскелета, в гидрогеносомах и пероксисомах — специальных вакуолях, осуществляющих реакции с выделением водорода и утилизацию перекиси водорода.
С применением более совершенных методов эти открытия пришлось «закрыть», но оказалось, что пероксисомы и гидрогеносомы почему-то получают новые липиды для мембран через ту же систему транспортных белков, что и митохондрии. Тогда как нормальные вакуоли получают новые липиды в виде целых мембранных пузырьков.
Эта особенность пероксисом может означать, что они произошли от митохондрий путем крайнего упрощения и полной потери генома.
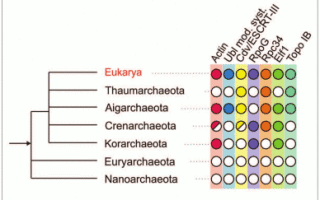
Гораздо сложнее было установить природу ядерно-цитоплазматического компонента (ЯЦК) эукариотической клетки. Сегодня, когда прочитаны последовательности геномов более чем тысячи видов бактерий и архей, появилась возможность методами сравнительной геномики искать прокариотные гомологи каждого из тысяч эукариотических белков. Краткие результаты такого поиска представлены на рисунке:
Из http://evolbiol.ru/dok_ibr2009.htm
Сразу видно, что базовые генетические процессы эукариотических клеток — репликация, транскрипция, трансляция, репарация — обслуживаются белками практически строго архейного происхождения.
Гены этих групп реже всего подвергаются горизонтальному переносу, и скорее всего были унаследованы эукариотами напрямую.
ДНК- и РНК-полимеразы бактериального происхождения, кодируемые эукариотическими геномами, работают в митохондриях и пластидах и явно перенесены из их геномов. Это значит, что предок ЯЦК, скорее всего, принадлежал к археям.
Происхождение митохондрий благодаря сравнительной геномике удалось сильно уточнить. Их бактериальные предки относились к группе альфа-протеобактерий, причем в современном мире к ним ближе всего находятся пурпурные бактерии Rhodospirillum.
Эти бактерии способны как к фотосинтезу, используя соединения серы для восстановления CO2, так и к аэробному дыханию на готовой органике, переключаясь между этими режимами метаболизма в зависимости от наличия кислорода и света.
Большая часть их дыхательной цепи работает как в фотосинтезе, так и в аэробном дыхании.
Ранее считалось, что некоторые безмитохондриальные эукариоты (Giardia, микроспоридии) отделились от остальных в глубокой древности и никогда не имели митохондрий. Сегодня выясняется, что в их ядерном геноме есть гены альфа-протеобактериального происхождения, что свидетельствует о потере когда-то имевшихся митохондрий.
Одна из малоизвестных функций митохондрий — сборка железо-серных кластеров ферментов, в том числе, работающих в цитоплазме. Для этого требуется транспорт свернутых белковых глобул, содержащих железо-серные кластеры, через митохондриальную мембрану.
Это достаточно сложное и затратное приспособление, которое могло возникнуть только если хозяин митохондрий сам не умел собирать железо-серные кластеры.
Простейшее предположение о природе белков бактериального происхождения, работающих в цитоплазме эукариот — что они все принадлежали предку митохондрий или пластид — оказывается неверным.
У эукариот насчитывается более 200 генных семейств бактериального происхождения, отсутствующих у альфа-протеобактерий и цианобактерий, и следовательно, приобретенных другими путями. Другие белки эукариот, хотя имеют альфа-протеобактериальные гомологи, ближе к белкам других групп бактерий.
Например, все ферменты гликолиза эукариот ближе всего к белкам бактерий-бродильщиков рода Clostridium.
Из http://evolbiol.ru/dok_ibr2009.htm
Несколько белковых семейств эукариот, связанных со слиянием и отшнуровкой мембран, имеют архейное происхождение, например, Adaptin N, Syntaxin, AdAR repeat. К сожалению, их функции в клетках архей неизвестны.
Помимо митохондрий, для приспособления к кислороду нужны белки, защищающие цитоплазму, такие как оксигеназы, пероксидазы, каталаза.
Эти белки эукариот все получены от бактерий, причем, не родственных митохондриям.
Этого и следовало ожидать — ведь чтобы получать выгоду от аэробного дыхания, надо сначала научиться защищаться от токсического действия кислорода, которое проявляется уже в очень малых концентрациях.
Мембраны эукариот, подобно бактериальным, состоят из сложных эфиров жирных кислот. Ферменты биосинтеза липидов, соответственно, могли быть получены только от бактерий. Уникальные липиды эукариот — стеролы, такие как холестерол, синтезируются из терпеновых предшественников.
Ключевой фермент их биосинтеза, сквален-монооксигеназа, относится к обширному семейству, общему для всех трех доменов жизни, но ближе всего к монооксигеназам актинобактерий. Сквален-синтаза эукариот одинаково близка и к бактериальным, и к архейным сквален-синтазам.
Холестерол придает мембранам подвижность и текучесть, что облегчает фагоцитоз, и его биосинтез, скорее всего, был необходим для перехода к фагоцитозу и приобретению митохондрий.
Большая часть эукариотических генов бактериального происхождения, не принадлежавших предкам митохондрий и пластид, кодирует защитные, рецепторные, транспортные, сигнальные и регуляторные белки.
Сюда относятся, например, белки теплового шока Hsp90, белки TerC и Ttg2, связанные с устойчивостью к тяжелым металлам, семейство Ras, передающее сигналы в протеин-киназных путях, и другие.
Что интересно, белки, которые у эукариот обеспечивают взаимодействия между компонентами клетки, у бактерий нужны для связей между разными клеткам в сообществе.
Например, Tim44 эукариот нужен для транспорта белков через внутреннюю мембрану митохондрий из цитоплазмы, а его бактериальные гомологи экспортируют белки из клеток во внешнюю среду. Это свидетельствует о том, что при возникновении эукариотической клетки под одной мембраной оказались уже отработанные системы связи клеток в сообществе.
Происхождение цитоскелетных белков прояснилось буквально в последние три года. Ранее были известны белки FtsZ, отдаленно похожие по последовательности на тубулин, широко распространенные у бактерий и архей, полимеризующиеся в пластины и трубки, и необходимые для деления прокариотических клеток.
В 2012 году были найдены артубулины — близкие гомологи тубулина – у двух видов архей рода Nitrosoarchaeum, принадлежащих к группе Thaumarchaeota . Их функция пока неизвестна.
Аналогично, у бактерий и архей широко распространены отдаленные гомологи актина — белки MreB, полимеризующиеся в тонкие нити и участвующие в перетяжке делящейся бактериальной клетки. Другие дальние гомологи актина кодируются плазмидами и служат для их расхождения при делении клетки.
И только в 2009 году у нескольких видов группы Crenarchaeota были обнаружены кренактины — близкие гомологи эукариотического актина. Кренактины образуют нити, которые придают клеткам палочковидную форму, часто с разветвлениями.
По своей последовательности кренактины близки не только к актинам, но и белкам Arp2/Arp3 (Actin-related proteins), которые у эукариот образуют разветвления актиновых филаментов. В некоторых случаях полимеризация актина с Arp2/Arp3 на плазматической мебране достаточна для отшнуровки фагоцитозного пузырька.
Наряду с артублинами у Thaumarchaeota обнаружились гомологи еще одной эукариотической системы — ESCRT-III (endosomal sorting complex required for transport).
Эта группа белков связана с мембранами комплекса Гольджи, эндоплазматического ретикулума и вакуолей и участвует в сортировке содержимого мембранных пузырьков.
В клетках архей белки этого комплекса необходимы для деления клеток, заменяя отсутствующий FtsZ.
В геноме Caldiarchaeum subterranum, прочитанном в 2010 году, были обнаружены гомологи эукариотической системы убиквитиновой модификации белков.
Эта система пришивает убиквитин — маленький белок — к различным клеточным белкам, что служит обычно меткой для их уничтожения в протеосоме, а так же влияет на другие процессы, например, перестройку хроматина.
Кроме собственно убиквитина (Ub), в нее в минимальном варианте входят три фермента — Е1, Е2 и Е3 — катализирующие разные стадии присоединения Ub. У бактерий известны отдаленные гомологи Е1 и Е2, но их функции не связаны с убиквитином, они катализируют включение серы при синтезе витамина В1 и молибденовых кластеров ферментов.
Протеосомы были известны ранее у некоторых бактерий (Mycobacterium tuberculosum) и архей (Haloferax volcanii), но убиквитиновой системы у этих видов нет. Протеосомы есть и у Caldiarchaeum subterranum, они похожи на эукариотические также по наличию белка RPN11, который опознает убиквитиновые метки на белках.
Гомологи различных эукариотических систем рассеяны по разным группам архей. Ubi — убиквитиновая система модификации белков; ESCRT-III — система деления клетки (у архей) и система сортировки мембранных пузырьков (у эукариот); RpoG и Rpc34 — вспомогательные субъединицы РНК-полимераз; Elf1 – фактор элонгации транскрипции; TopoIB — топоизомераза IB. (Bernander et al., 2011)
Таким образом, мы видим, что ЯЦК эукариотической клетки сочетает в себе черты архей и различных неродственных групп бактерий.
От архей унаследованы центральные информационные системы клетки, зачатки цитоскелета, зачатки систем управления мембранами и убиквитиновая система модификации белков.
От бактерий произошли ферменты метаболизма сахаров, липидов и отчасти – стеролов, системы защиты от кислорода и всевозможные сигнальные и регуляторные белки.
Очевидно, что приобретение митохондрий было далеко не первым шагом на пути происхождения эукариот: чтобы такой симбиоз стал механически возможен, необходимы механизмы фагоцитоза. А чтобы он был еще и полезен, надо сначала иметь ферменты гликолиза, защиту цитоплазмы от кислорода, системы транспорта и передачи сигналов между симбионтом и хозяином.
https://www.youtube.com/watch?v=2tqs40MUsJc\u0026t=995s
Как мог происходить процесс появления эукариот? Во-первых, симбиогенез, многочисленные переносы генов от разных бактерий и большая роль сигнально-регуляторных генов означают, что процесс происходил в сложном сообществе прокариот.
Во-вторых, архейное происхождение базовых генетических систем эукариот свидетельствует о том, что у истоков процесса стояли археи. Из разнообразных экологических групп современных архей только одна, метаногены, живет в сложных сообществах в тесном взаимодействии с бактериями.
Остальные археи, как правило, специализированны к экстремальным местообитаниям, где образуют крайне бедные сообщества с малой ролью межвидовых взаимодействий. Отсутствие всяких следов метаногенеза у эукариот означает, что этот тип метаболизма в какой-то момент эукариогенеза стал непригоден и был заменен другими.
В качестве причины кризиса метаногенов напрашивается появление кислородного фотосинтеза. Ферменты метаногенеза используют ионы двухвалентного никеля, которые очень уязвимы для кислорода.
Итак, скорее всего, эукариогенез начался с метаногенной археи, обитавшей в нижних слоях бактериального мата в симбиозе с ацетогенными бродильщиками, выделяющими водород. Появление кислородного фотосинтеза в верхнем слое того же мата вызвало крупный экологический кризис.
Многие обитатели мата вымерли, другим же удалось создать свои или приобрести горизонтальным переносом чужие системы защиты от кислорода. Этот процесс шел не одновременно, а распространялся с поверхности в глубокие слои мата, и к моменту появления серьезных проблем у метаногенов их соседи сверху уже имели адаптации к кислороду.
Многие прокариоты в состоянии стресса начинают активно поглощать ДНК из внешней среды, и таким способом наша архея приобрела защиту от кислорода и новый метаболизм, скорее всего молочнокислое брожение. Сквален-монооксигеназа, необходимая для синтеза стеролов, могла исходно служить для защиты от кислорода.
Отсутствие клеточной стенки, актиновый цитоскелет для поддержания вытянутой и ветвистой формы клеток, стеролы и освобождение мембраны от энергетических функций позволили ей перейти к фагоцитозу и успешно конкурировать с соседями-бактериями.
Такой способ питания усилил поток чужеродных генов в цитоплазму клетки, что, видимо, потребовало специализации трех РНК-полимераз из одной предковой.
Тем временем в среднем слое мата, среди пурпурных фотосинтезирующих бактерий, было отработано аэробное дыхание, для которого ценным субстратом стали органические кислоты, выделяемые бродильщиками нижних слоев. Эти аэробы стали оптимальными партнерами для симбиоза с фагоцитирующими бродильщиками. Вероятно, сначала взаимодействие между ними было отработано на стадии простого соседства в мате, а затем продолжилось с переходом к эндосимбиозу.
Поток чужеродных генов неизбежно должен был заражать фаготрофную клетку новыми видами вирусов и мобильных элементов. Их давление могло вызвать ответ в виде образования ядра для защиты генетического материала.
Разделение клетки на компартменты, один из которых специализируется на хранении ДНК, известно и у бактерий группы Planctomycetes (к сожалению, чтение и аннотация их геномов пока не закончена).
Появление ядра, разделившее трансляцию и транскрипцию, сделала возможным размножение интронов в геноме, впоследствии из них спецализировалась сплайсеома. Новые мобильные элементы резко активизировали рекомбинацию, что могло привести к распаду кольцевой ДНК на линейные хромосомы.
Функция защиты их концов могла сначала выполняться мобильными элементами, размножающимися параллельно делению клетки, подобно тому как это происходит у ряда насекомых, затем на основе обратной транскриптазы ретротранспозонов возникла теломераза.
После приобретения митохондрий протоэукариоты стали высокоэффективными хищниками: аэробное дыхание повышает эффективность использования пищи почти в 20 раз по сравнению с гликолизом.
Они смогли выйти на поверхность мата и фагоцитировать цианобактерий.
Так часть эукариот приобрели пластиды, смогли оторваться от бактериального мата и перейти к жизни в толще океана, другие же остались гетеротрофами.
(раздел слишком велик для одного поста – продолжение следует)
Происхождение эукариот
☰
Расцвет эукариот на Земле начался около 1 млрд лет назад, хотя первые из них появились намного раньше (возможно 2,5 млрд лет назад). Происхождение эукариот могло быть связано с вынужденной эволюцией прокариотических организмов в атмосфере, которая стала содержать кислород.
Симбиогенез — основная гипотеза происхождения эукариот
Существует несколько гипотез о путях возникновения эукариотических клеток. Наиболее популярная — симбиотическая гипотеза (симбиогенез).
Согласно ей, эукариоты произошли в результате объединения в одной клетке разных прокариот, которые сначала вступили в симбиоз, а затем, все более специализируясь, стали органоидами единого организма-клетки.
Как минимум симбиотическое происхождение имеют митохондрии и хлоропласты (пластиды вообще). Произошли они от бактериальных симбионтов.
Клеткой-хозяином мог быть относительно крупный анаэробный гетеротрофный прокариот, похожий на амебу. В отличие от других, он мог приобрести способность питаться путем фаго- и пиноцитоза, что позволяло ему захватывать других прокариот. Они не все переваривались, а снабжали хозяина продуктами своей жизнедеятельности). В свою очередь, получали от него питательные вещества.
Митохондрии произошли от аэробных бактерий и позволили клетке-хозяину перейти к аэробному дыханию, которое не только намного эффективней, но и облегчает существование в атмосфере, содержащей достаточно большое количество кислорода. В такой среде аэробные организмы получают преимущество над анаэробными.
Позже в некоторых клетках поселились похожие на ныне живущих синезеленых водорослей (цианобактерий) древние прокариоты. Они стали хлоропластами, дав начало эволюционной ветви растений.
Кроме митохондрий и пластид симбиотическое происхождение могут иметь жгутики эукариот. В них превратились симбионты-бактерии наподобие современных спирохет, имеющих жгутик. Считается, что в последствии из базальных тел жгутиков произошли центриоли, столь важные структуры для механизма клеточного деления эукариот.
Эндоплазматическая сеть, комплекс Гольджи, пузырьки и вакуоли могли произойти от наружной мембраны ядерной оболочки. С другой точки зрения, некоторые из перечисленных органелл могли возникнуть путем упрощения митохондрий или пластид.
Во многом неясным остается вопрос происхождения ядра. Могло ли оно также образоваться из прокариота-симбионта? Количество ДНК в ядре современных эукариот во много раз превышает его количество в митохондриях и хлоропластах. Возможно часть генетической информации последних со временем переместилась в ядро. Также в процессе эволюции происходило дальнейшее увеличение размера ядерного генома.
Кроме того в симбиотической гипотезе происхождения эукариот не все так однозначно с клеткой-хозяином. Им мог и не быть один вид прокариот. Используя методы сравнения геномов, ученые делают вывод, что клетка-хозяин близок к археям, при этом сочетает в себе признаки архей и ряда неродственных групп бактерий.
Отсюда можно сделать вывод, что появление эукариот происходило в сложном сообществе прокариот. При этом процесс скорее всего начался с метаногенной археи, вступавшей в симбиоз с другими прокариотами, что было вызвано необходимостью обитания в кислородной среде.
Появление фагоцитоза способствовало притоку чужих генов, а ядро образовалось для защиты генетического материала.
Молекулярный анализ показал, что различные белки эукариот происходят от разных групп прокариот.
Доказательства симбиогенеза
В пользу симбиотического происхождения эукариот говорит то, что митохондрии и хлоропласты имеют собственную ДНК, причем кольцевую и не связанную с белками (также обстоит дело у прокариот). Однако в генах митохондрий и пластид есть интроны, чего нет у прокариот.
Пластиды и митохондрии не воспроизводятся клеткой с нуля. Они образуются из ранее существующих таких же органелл путем их деления и последующего роста.
В настоящее время существуют амебы, у которых нет митохондрий, а вместо них есть бактерии симбионты. Также есть простейшие, сожительствующие с одноклеточными водорослями, выполняющими в клетке-хозяине роль хлоропластов.
Инвагинационная гипотеза происхождения эукариот
Кроме симбиогенеза существуют и другие взгляды на происхождение эукариот. Например, инвагинационная гипотеза. Согласно ей, предком эукариотической клетки был не анаэробный, а аэробный прокариот. К такой клетке могли прикрепляться другие прокариоты. Потом их геномы объединялись.
Ядро, митохондрии и пластиды возникли путем впячивания и отшнуровывания участков клеточной мембраны. В эти структуры попадала чужеродная ДНК.
Усложнение генома происходило в процессе дальнейшей эволюции.
Инвагинационная гипотеза происхождения эукариот хорошо объясняет наличие двойной мембраны у органелл. Однако она не объясняет, почему система биосинтеза белка в хлоропластах и митохондриях сходна с прокариотической, в то время как таковая в ядерно-цитоплазматическом комплексе имеет ключевые отличия.
Причины эволюции эукариот
Все разнообразие жизни на Земле (от простейших до покрытосеменных и млекопитающих) дали клетки эукариотического, а не прокариотического типа. Возникает вопрос, почему? Очевидно, ряд особенностей, возникших у эукариот, существенно повысили их эволюционные возможности.
Во-первых, у эукариот есть ядерный геном, который во много раз превосходит количество ДНК у прокариот. При этом эукариотические клетки диплоидны, кроме этого в каждом гаплоидном наборе определенные гены многократно повторяются.
Все это обеспечивает, с одной стороны, большие масштабы для мутационной изменчивости, а с другой — уменьшает угрозу резкого снижения жизнеспособности в результате вредной мутации.
Таким образом, эукариоты, в отличие от прокариот, обладают резервом наследственной изменчивости.
Эукариотические клетки имеют более сложный механизм регуляции жизнедеятельности, у них существенно больше различных регуляторных генов. Кроме того, молекулы ДНК образовали комплексы с белками, что позволило наследственному материалу упаковываться и распаковываться.
Все вместе это дало возможность считывать информацию частями, в разных сочетаниях и количестве, в разное время. (Если в клетках прокариот транскрибируется почти вся информация генома, то в эукариотических клетках обычно менее половины.
) Благодаря этому эукариоты могли специализироваться, лучше приспосабливаться.
У эукариот появились митоз, а затем и мейоз. Митоз позволяет воспроизводить генетически сходные клетки, а мейоз сильно увеличивает комбинативную изменчивость, что ускоряет эволюцию.
Большую роль в процветании эукариот сыграло приобретенное их предком аэробное дыхание (хотя оно есть и у многих прокариот).
На заре своей эволюции эукариоты обзавелись эластичной оболочкой, обеспечивавшей возможность фагоцитоза, и жгутиками, позволившими им двигаться. Это дало возможность эффективней питаться.
plustilino © 2019. All Rights Reserved
Непростые пути эволюции. Откуда взялись эукариоты?
Земля существует около 4,5 млрд. лет. Большую часть этого времени её население составляли исключительно одноклеточные. Попытки создать нечто большее, объединившись в сообщество, предпринимались очень рано, окаменелостям древнейших бактериальных матов много больше трёх миллиардов лет.
Но попытки были не слишком удачными, сообщества так и оставались всего лишь сообществами, не превратившись в полноценный многоклеточный организм. Так продолжалось до тех пор, пока на Земле не появились эукариоты. Возникновение эукариотической клетки стало огромным скачком в развитии земной жизни и подготовило следующий этап – появление многоклеточных.
До самого последнего времени о том, как это произошло, было известно очень мало.
Амёба. Типичный представитель одноклеточных эукариотАмёба. Типичный представитель одноклеточных эукариот
Ещё совсем недавно мы учили в школе, что живые существа делятся на два больших надцарства: прокариоты, клетка которых не имеет оформленного ядра, и эукариоты, чья клетка оформленное ядро имеет.
Правда, есть ещё вирусы, которые обходятся вовсе без клеток, но это – очень отдельный случай, и непонятно даже, стоит ли причислять их к живым организмам.
К прокариотам относятся бактерии (Bacteria), к эукариотам – Простейшие (Protozoa), водоросли (кроме сине-зелёных, которые цианобактерии) и все виды многоклеточных организмов.
Предполагалось, что сначала бактерии эволюционировали до эукариот, после чего смогли совершить ещё один глобальный эволюционный прорыв, который никак не давался бактериям – перейти к настоящей многоклеточности. Притом, слова «прокариоты» и «бактерии» употреблялись практически как синонимы. Однако в действительности всё оказалось несколько сложнее.
Отличия прокариотической и эукариотической клетокОтличия прокариотической и эукариотической клеток
Надо признать, люди довольно мало знают о прокариотах. Связано это с тем, что как следует изучить их можно лишь в лаборатории, а далеко не все из них получается разводить в лабораторных условиях.
Долгое время изучались исключительно культивируемые виды, что сродни сюжету известного анекдота – поиску потерянного кошелька под фонарём. Некультивируемых прокариот гораздо больше чем культивируемых.
Не приходится удивляться, что человечество очень долго оставалось в неведении относительно того факта, что далеко не все прокариоты являются бактериями.
Открытия, сделанные в 70-е годы XX века полностью перевернули старые представления о классификации одноклеточных.
Произошло это в ходе изучения метаногенных бактерий-экстремофилов, живущих в слишком горячей или слишком солёной для большинства организмов воде и выделяющих в качестве продукта жизнедеятельности метан.
По понятным причинам, подробности их устройства всегда вызывали особенно сильное любопытство. И вот оказалось, что некоторые из этих «бактерий» устроены настолько необычно, что их и бактериями-то не назовёшь и в родстве с ранее известными бактериями не заподозришь.
И это при огромном внешнем сходстве! Размер клетки, отсутствие оформленного ядра и мембранных органелл, всё как у людей бактерий, но биохимия имеет особенности, указывающие на то, что бактерии и новооткрытые незнакомцы возникли и развивались абсолютно независимо друг от друга!
Незнакомцы получили название археи, а их первооткрыватель, американский микробиолог Карл Ричард Вёзе ввёл в употребление трёхдоменную систему классификации. Отныне живые организмы надлежало делить не на два, а на три большие домена: бактерии, археи и эукариоты.
Карл Ричард Вёзе
Поначалу решили, что археи – очень экзотические организмы, живущие в очень экзотических условиях.
Но когда их стали искать целенаправленно, то выяснилось, что, во-первых, экстремофилов полно и среди настоящих бактерий, и во-вторых, археи встречаются отнюдь не только в горячих вулканических источниках, а практически повсюду: в почве, воде, воздухе, кишечнике высших животных.
Возможно, планктонные археи — самая многочисленная группа ныне живущих организмов. Просто раньше они не попадали в поле зрения. Их считали бактериями и не изучали пристально. Во-первых, потому что плохо культивируются, во-вторых, потому что в отличие от бактерий для человека безвредны.
До сих пор не обнаружено ни одного патогенного (болезнетворного) вида. Правда, обитающие в пищеварительном тракте человека и жвачных животных археи осуществляют полезную функцию, помогая пищеварению. Но, как говорила старуха Шапокляк, хорошими делами не прославишься.
Итак, исходя из особенностей клетки, всё живое делится на три больших домена: бактерии, археи и эукариоты. Причём первые и вторые друг от друга не происходят.
А вот от кого происходят эукариоты, это оказалась задача не на одну трубку. Между тем, она представляет исключительный интерес. Появление эукариотической клетки ( приблизительно 1,6 – 2,1 млрд.
лет назад) – важнейший момент биологической эволюции.
Бактериальные сообщества никогда не переходят грань отделяющую колонию от единого организма.
Составляющие его клетки не меняются настолько, чтобы колония не могла распасться, а её обитатели вернуться к одиночному образу жизни.
Появлению многоклеточных предшествовало ещё одно глобальное изменение – возникновение эукариотической клетки, способной в дальнейшем развиваться столь разнообразно, что это привело к дифференциации тканей живых организмов.
Помимо наличия оформленного ядра эукариоты отличаются от бактерий и архей и другими признаками. Самый заметный из них – размер. Прокариоты имеют размеры порядка 1 мкм, эукариотическая клетка от 10 до 100 мкм. То есть, с появлением эукариот линейный размер живых существ на Земле увеличился на порядок. И это означает разницу в объёме в тысячи раз.
Ещё одним важнейшим отличием является наличие органелл, особых структур, расположенных в цитоплазме эукариотической клетки.
Примером органелл могут служить, например митохондрии , которые выполняют функцию энергетической станции клетки.
В них происходит окисление органических соединений и использование освобождающейся при их распаде энергии для генерации электрического потенциала, синтеза аденозинтрифосфорной кислоты (АТФ) и термогенеза.
Инфузории-туфельки, одноклеточные эукариотыИнфузории-туфельки, одноклеточные эукариоты
Собственно, клетка с органеллами – черновой вариант многоклеточного существа. Когда-то предки органелл были свободными прокариотами и гуляли сами по себе, но были захвачены посторонней клеткой и поставлены ей на службу. В спорах о том, как именно это происходило, сломали много копий.
Самой популярной была теория фагоцитоза, согласно которой одна клетка попросту глотала другую, но не переваривала , а встраивала полезные в хозяйстве элементы в собственный организм. Сейчас в эту теорию вносят некоторые поправки. Но мы забегаем вперёд.
Ряд важных отличий эукариот от бактерий лежит в биохимической области. С открытием архей выяснилось, что во многих отношениях эукариоты как раз ближе к археям. В связи с этим встал вопрос, а не являются ли как раз археи, а вовсе не бактерии прямыми предками эукариот? Или быть может археи и эукариоты произошли от одного общего предка?
Изучение этого вопроса сопряжено с большими трудностями, как и вообще всё, что касается архей. Как уже говорилось, археи не культивируются в лабораторных условиях. Для того, чтобы узнать о них больше используется так называемый метагеномный анализ.
Из среды обитания, скажем из придонных отложений, извлекают обрывки ДНК и после тщательного изучения и сопоставления данных моделируют то, как это выглядело, когда было целым. Так учёные узнают о существовании новых видов микроорганизмов и ищут среди них тех, что могли бы быть близкими родственниками эукариот.
В чём-то это сродни палеонтологии, когда найдя одну косточку тут, другую там со временем проводят реконструкцию облика ранее неизвестного науке животного. Только восстанавливают из фрагментов не скелет, а геном.
На сегодняшний день установлено, что так называемый ядерно-цитоплазматический компонент эукариотической клетки находится в близком родстве с археями, её же митохондрии и пластиды являются потомками захваченных «хозяйской» клеткой бактерий (альфапротеобактерий и цианобактерий соответственно).
Что до характера родства эукариот и архей, тот тут возможны вопросы. Эукариоты либо являются потомками архей, либо происходят от общего с ними предка. Во втором случае система классификации остаётся по-прежнему трёхдоменной.
Если же верно первое, то её надо снова менять на двухдоменную, только на этот раз живые организмы делятся на два больших домена: бактерии и археи, а последние включают в себя эукариот.
Чтобы прояснить ситуацию, нужно было найти «недостающее звено», архею, максимально близкую генетически к эукариотам. В идеале более близкую чем к другим археям. И такое открытие было сделано.
В 2015 г. исследователи из Уппсальского университета (Швеция) и Бергенского университета (Норвегия) изучили ДНК из пробы грунта, поднятой с глубины 3283 м в Северном Ледовитом океане, недалеко от гидротермальной зоны Loki’s Castle («Замок Локи»), расположенной на склоне срединно-океанического хребта.
Им удалось собрать из кусочков почти полный геном ранее неизвестного науке организма, относящегося к глубоководной группе архей. После публикации исследования группа получила название локиархеи (Lokiarchaeota).
Кроме восстановленного почти полностью генома удалось частично восстановить геномы ещё двух организмов этой группы, получивших названия Loki2 и Loki3.
Когда, исходя из новых генетических данных стали строить эволюционное древо, выяснилось, что эукариоты не просто принадлежат к древу архей, а находятся не у его корня, а довольно высоко в ветвящейся кроне. Дальнейший метагеномный анализ глубоководных остатков показал, что локиархени входят в большой надтип включающий минимум четыре веточки. Всю эту большую ветвь назвали асгарархеями.
Они действительно генетически ближе к эукариотам, чем к другим ветвям архей. В их геноме даже были закодированы белки, которые до сих пор считались уникальными для эукариот.
Особенный интерес представляют гены, связанные с подвижностью клетки и ее мембраны, с возможностью формирования разнообразных мембранных структур и активного захвата объектов из внешней среды.
Их изучение может пролить свет на увлекательную историю приобретения эукариотами пластид и митохондрий.
Но геном – это только геном. Безусловно, он даёт учёным некоторую информацию о том, как выглядит весь организм, но на нынешнем уровне развития генетики далеко не полную. Несколько лет заветной мечтой микробиологов оставалось получить в своё распоряжение живые асгардархеи. И вот, наконец, эта мечта сбылась.
“Синкай-6500”
В августе 2019 г. была опубликована научная работа об исследовании донного грунта, который японский батискаф «Синкай-6500» извлёк с глубины 2,5 км.
Пробы поместили в биореактор, а спустя какое-то время взяли из него вторичные пробы, предположительно обогащенные живыми клетками, и перенесли их на питательные среды.
В ходе изучения того, что получилось, обнаружили микроорганизм, получивший название Prometheoarchaeum syntrophicum. Тщательнейший молекулярно-генетический анализ показал, что он относится к группе локиархей.
Прометеоархеум под электронным микроскопом. Прометеоархеум под электронным микроскопом.
Прометеоархеум имеет простую сферическую форму. Диаметр описанных клеток — от 300 до 750 нанометров, а в среднем 550 нанометров, что приблизительно соответствует нижней границе характерного для прокариот размера.
Обычно клетки прометеоархеума образуют цепочки, окруженные полисахаридными чехлами. Электронная микроскопия показала, что в клетках прометеоархеума нет никаких внутренних включений, напоминающих эукариотные органеллы. «Начинка» клетки у них совершенно прокариотная.
Зато в клетке есть хорошо отрегулированная система перестройки мембран, и это очень важный момент.
Давно существовала теория, что предшественники эукариот «научились» создавать сначала выросты клетки, и только потом впячивания. Это разные процессы, обслуживаемые разными белками, причем те сопутствующие белки цитоскелета, которые нужны для впячиваний, появились позже, чем те, которые нужны для выростов.
Теория подтвердилась. Оказалось, что клетки прометеоархеума регулярно образуют длинные выросты, иногда прямые, но часто сложным образом ветвящиеся, а на впячивания они не способны.
Между тем, без этого невозможен фагоцитоз и, скорее всего, потомки локиархей приобрели органеллы другим способом. Это и раньше подозревали.
Фагоцитоз энергоемок и на промежуточном этапе, до возникновения полноценных энергетических батарей митахондрий мог привести не к выигрышу, а к проигрышу.
Для того, чтобы понять, как всё-таки произошёл переход к эукариотической клетке следует присмотреться к метаболизму новооткрытой археи и к характерным для неё условиям жизни. Прометеоархеум — анаэроб, ему не требуется кислород. Питательными веществами ему служат аминокислоты — аланин, серин, цистеин, аспартат, триптофан, тирозин, гистидин, треонин, лизин.
Молекулы аминокислот в клетке прометеоархеума подвергаются бескислородному окислению, среди конечных продуктов которого после длинной цепочки реакций оказываются водород (H2) и муравьиная кислота, она же формиат. Есть также метаболические цепочки, выдающие на выходе уклекислоту.
А живут прометеоархеумы в тесном соседстве с двумя другими группами микробов: метаногенными археями и сульфатредуцирующими бактериями. Метаногены и сульфатредукторы получают от прометеоархеума конечные продукты обмена веществ: формиат, углекислоту и водород. Прометеоархеуму отработанный водород больше не нужен, зато он необходим метаногенам и сульфатредукторам.
Взамен и метаногены, и сульфатредукторы поставляют прометеоархеуму аминокислоты, а также витамины и другие полезные вещества, которые сам он синтезировать не умеет. Такая химическая взаимозависимость микробов называется синтрофией. Она не требует дополнительных затрат. До поры до времени одноклеточные просто живут рядом и пользуются услугами друг друга.
Притом, синтрофия никак не зависит от родства: известны случаи синтрофных отношений между бактериями и археями, то есть того, что в принципе и нужно для происхождения эукариот.
Если клетка археи образует для увеличения поверхности мощные выросты, симбиотическая бактерия может оказаться между ними и продолжать прежний образ жизни. Затем выросты ветвятся, сливаются и бактерия оказывается заперта в полости, где её неплохо кормят.
Со временем система замкнутых полостей стала эндоплазматической сетью эукариотической клетки. А митохондрии пробуравили мембрану, ограничивающую эти полости, и выбрались прямо в цитоплазму. Так возникла удивительная химера, объединяющая в себе признаки совершенно разных организмов: археи и бактерии.
И она стала стартовой площадкой для следующего глобального рывка эволюции.