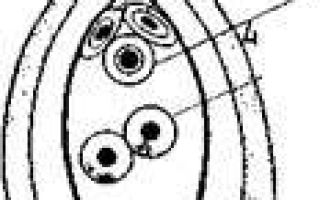
Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап — спорогенез — завершается образованием гаплоидных клеток — спор, в ходе 2-го этапа — гаметогенеза — происходит ряд делений гаплоидных клеток, прежде чем образуются зрелые гаметы.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) — мега- или макроспорогенезом. Микроспорогенез протекает аналогично делению созревания у животных мужских половых клеток до стадии сперматиды, а мегаспорогенез — соответственно до стадии незрелой яйцеклетки — ооцита II.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У животных после двух мейотических делений формируются гаметы, и никаких дополнительных клеточных делений не происходит.
У растений в результате двух мейотических делений возникает гаплоидная спора, из которой развивается гаметофит, представляющий собой у низших растений (грибов, печеночников, мхов, ряда водорослей) целый организм и наиболее продолжительную стадию цикла существования.
У высших растений гаплоидная фаза редуцирована, однако ядра мужской и женской спор претерпевают ряд митотических делений, прежде чем образуются гаметы.
Микроспорогенез и микрогаметогенез
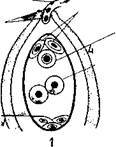
Мы рассмотрим микроспорогенез и микрогаметогенез на примере покрытосеменных растений как наиболее общем. В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делении становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
Схема микроспорогенеза (1-6) и микрогаметогенеза (5-10) у растений
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
У однодольных растений каждое деление ядра в мейозе, как правило, сопровождается цитокинезом; у двудольных оба деления клетки наступают одновременно по окончании мейоза.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, кутинизированная, поверхность ее либо гладкая, либо шероховатая; приспособленная для переноса пыльцы и прилипания ее к рыльцу пестика.
Этим заканчивается микроспорогенез вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся.
В ней накапливаются запасные питательные вещества, который в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые в отличие от сперматозоидов животных называются спермиоклетками, или спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: Два из них — спермии и одно — вегетативное. При образовании пыльцевой трубки это вегетативное ядро в полужидком диффузном состоянии переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены С. Г. Навашиным в 1910 г. на лилейных растениях.
Мегаспорогенез и мегагаметогенез
У покрытосеменных растений женский гаметофит — это зародышевый мешок, который закладывается и развивается внутри семяпочки.
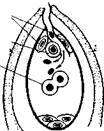
Схема мегаспорогенеза (1-5) и мегагаметогенеза (6-12) у растений
Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез. В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоры.
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной.
Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития), судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем ее ядро претерпевает ряд эквационных делений. При этом сама клетка не делится, делится только ядро.
У разных систематических групп растений число эквационных делений ядра мегаспоры может варьировать от одного до трех.
У большинства растений (70% видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других — в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка, и две так называемые синергиды образуют яйцевой аппарат. Однако из этих трех клеток после оплодотворения развивается только одна, а две другие разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца.
Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное или центральное, ядро зародышевого мешка. Это ядро с цитоплазмой зародышевого мешка называют обычно центральной клеткой зародышевого мешка. Однако часто полярные ядра, передвинувшиеся к центру, не сливаются до оплодотворения.
Оставшиеся у халазального конца зародышевого мешка три ядра также обособляются в клетки; они называются антиподами.
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 наследственно одинаковых гаплоидных ядер, из которых только одно дает яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно.
В одних случаях, как мы только что рассмотрели, развитие зародышевого мешка начинается из одной гаплоидной споры (моноспорический тип развития), в других — из двух (биспорический тип) и четырех спор (тетраспорический тип).
Типы развития зародышевых мешков
Как мы указывали, при моноспорическом типе развивается лишь одна мегаспора из четырех, а остальные три разрушаются подобно тому, что имеет место с редукционными тельцами у животных. При других типах развития зародышевого мешка сохраняется разное количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям.
Образование пыльцевых зерен и зародышевых мешков у цветковых растений
Изучая гаметогенез, нельзя не поражаться тому параллелизму, который наблюдается при созревании половых клеток у животных и растений, несмотря на то, что их расхождение (дивергенция) в филогенезе произошло на очень раннем этапе возникновения клеточной организации. Это указывает на однотипность принципов построения ряда приспособительных механизмов как в растительном, так и животном мире.
Итак, изучение развития половых клеток у животных и у растений показало, что формирование гамет является сложным процессом. Прежде чем яйцеклетка и спермий объединятся в процессе оплодотворения, они претерпевают ряд превращений.
Однако половые клетки так же, как и клетки любой другой ткани, происходят из соматических. Поэтому их нельзя рассматривать как нечто обособленное от тела организма. Вместе с тем половые клетки имеют и свои особенности.
Основными характерными моментами, отличающими их от соматических клеток, являются следующие:
1. У разных животных и растений на разных стадиях дифференциации тканей зародыша происходит обособление половых клеток. Процесс закладки и дифференциации, половых клеток у животных называется зачатковым путем.
2. В процессе развития половых клеток особое значение имеет мейоз с характерными для него стадиями деления ядра, а именно профазой I, во время которой конъюгируют гомологичные хромосомы, метафазой I и анафазой I, когда осуществляется редукция числа хромосом и расхождение гомологичных хромосом к различным полюсам.
3. Главным свойством половых клеток является способность их при оплодотворении сливаться в одну с образованием зиготы, которая претерпевает затем дробление и развитие. Соматические клетки этой способностью, как правило, не обладают.
Спорогенез и гаметогенез у растений
Особенности гаметогенеза у растений. Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап — спорогенез — завершается образованием гаплоидных клеток — спор; 2-й этап — гаметогенез — включает образование зрелых гамет.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) —мега- или макроспорогенезом.
Микроспорогенез протекает аналогично сперматогенезу у животных, вплоть до образования сперматид, а мегаспорогенез соответствует образованию ооцита П.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У растений отсутствует зачатковый путь в том смысле, как это имеет место у животных.
Микроспорогенез и микрогаметогенез. Рассмотрим микроспорогенез и микрогаметогенез на наиболее общем примере покрытосеменных растений. В субэпидермальной ткани молодого пыльника, называемой археспорием, каждая клетка после ряда делений становится материнской клеткой пыльцы, которая проходит все фазы мейоза.
52
В результате двух мейотических делений возникают четыре гаплоидные микроспоры (рис. 24, /). Они лежат четверками, их называют тетрадами спор.
При созревании тетрады распадаются на отдельные микроспоры. Этим заканчивается микроспорогенез.
Вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток.
В дальнейшем вегетативная клетка и ее ядро не делятся.
В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, не способны к движению и называются спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три клетки. Две из них — спермин и одна вегетативная.
Мегаспорогенез и мегагаметогенез. В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, часто только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоры (см. рис._ 24, 2).
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады гаплоидна. Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития). Судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти, и затем ее ядро претерпевает ряд митозов. При этом сама клетка не делится, она образует зародышевый мешок.
У разных растений число митозов может варьировать от одного до трех. У большинства растений (70% видов покрытосеменных) бывает три деления, в результате возникает восемь одинаковых ядер.
Во время этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других в противоположном конце зародышевого мешка, называемого халазальным (рис. 25).
В дальнейшем ядра обособляются в самостоятельные клетки, имеющие значительное количество цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка и две так называемые синергиды — обра-
53
зуют яйцевой аппарат. Синергиды играют вспомогательную роль при оплодотворении, они скоро разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное, или центральное, ядро зародышевого мешка. Оставшиеся у халазального конца зародышевого мешка три ядра обособляются в клетки; они называются антиподами. Антиподы, так же как и синергиды, играют вспомогательную роль при развитии зиготы и вскоре разрушаются.
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 одинаковых гаплоидных ядер, из которых только одно образует яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно.
Кроме моноспорического типа развития, который только что рассмотрен, существуют и другие. При различных типах развития зародышевого мешка сохраняется разное'1 количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям (если сохраняются две мегаспоры — биспорический; четыре — тетраспорический тип развития).
Сравнение процессов созревания половых клеток у животных и растений (рис. 23 и 24) показывает почти полный параллелизм их, несмотря на то, что расхождение (дивергенция) растений и животных в филогенезе произошло на очень раннем этапе возникновения клеточной организации. Это указывает на однотипность принципов построения ряда приспособительных механизмов в растительном и животном мире.
ОПЛОДОТВОРЕНИЕ
Сущность оплодотворения. Оплодотворением принято называть побуждение яйца к развитию путем объединения в нем ядер (кариогамии) мужских и женских половых клеток.
Оплодотворение представляет собой необратимый процесс; однажды оплодотворенное яйцо не может быть оплодотворено вновь.
Син-гамия (слияние мужских и женских половых клеток) и кариогамия составляют сущность процесса оплодотворения.
Оплодотворение у животных. Процесс оплодотворения у животных можно разделить на несколько фаз. Первая фаза начинается с того, что сперматозоид либо прикрепляется к любой точке поверхности яйцеклетки, либо проникает в нее через микропиле. Момент соприкосновения головки сперматозоида с яйцом является начальным в цепи химических реакций. Эту фазу называют фазой активации яйца.
Вторая фаза процесса оплодотворения начинается после проникновения в яйцо одного (моноспермия), а у некоторых животных и нескольких сперматозоидов (полиспермия).
Проникший сперматозоид «готовится» к слиянию с женским ядром и последующему митозу: ядро сперматозоида постепенно набухает и приобретает вид интерфазного ядра. Такое ядро называют семенным или мужским пронуклеусом.
Ядро яйцеклетки, прошедшей все фазы мейоза, готовое к слиянию с ядром сперматозоида, называют женским пронуклеусом.
К моменту соприкосновения сперматозоида с яйцом и проникновения его внутрь ядро яйца у разных животных может находиться на разных стадиях деления созревания. Кариогамия же может произойти только после окончания мейоза, т. е. после образования женского пронуклеуса.
В акте оплодотворения два гаплоидных пронуклеуса сливаются в одно ядро, образуя зиготу. Этот момент является кульминационным пунктом процесса полового размножения. В результате кариогамии гомологичные хромосомы, разошедшиеся в мейозе у предыдущего поколения, вновь воссоединяются в одном ядре зиготы. Так восстанавливается диплоидный набор хромосом при половом размножении.
В цитоплазму яйцеклетки у млекопитающих проникает не только головка (ядро) сперматозоида, как это считали раньше, но и его шейка и даже хвостовая часть, что создает возможность передачи некоторого количества цитоплазмы мужского организма потомству.
Оплодотворение у растений. Оплодотворение у растений в принципе сходно с таковым у животных, но имеет и некоторые особенности.
Раньше было сказано, что микрогаметогенез завершается образованием двух спермиев, которые формируются или в пыльцевом зерне, или в пыльцевой трубке при прорастании пыльцевого зерна.
Пыльцевая трубка, дорастая до микропиле зародышевого мешка, соприкасается с яйцевым аппаратом — яйцеклеткой и синергидами (рис. 26, /). При соприкосновении конца пыльцевой трубки с синергидами пыльцевая трубка лопается, а синергиды разрушаются.
Передвигающиеся по пыльцевой трубке по мере ее роста два генеративных ядра — спермин после разрыва трубки вместе с ее содержимым попадают внутрь зародышевого мешка (рис. 26, 2). Из двух проникших в зародышевой мешок спермиев один спермин сливается с гаплоидным ядром яйцеклетки (рис. 26, 3).
Слияние ядра спермия с ядром яйцеклетки является собственно оплодотворением у растений. В оплодотворенной яйцеклетке — зиготе восстанавливается диплоидное число хромосом. Из зиготы развивается зародыш семени.
54
55
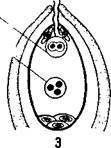
| Лыльиебая трудка ^ Синергиды Яйцеклетка |
| ВегетатиЬное ядро Спрр ' Мии |
| Зигота Триплоидное ядро шдоспврма |
|
Следует отметить, что у растений, так же как и у животных, женские ядра перед проникновением спермиев в зародышевый мешок могут находиться на разных стадиях, а кариогамия происходит лишь после завершения мейоза.
У покрытосеменных растений, кроме зародыша, в семени развивается дополнительный эмбриональный орган — эндосперм, который представляет собой питательное депо зародыша. Начало развития эндосперма обеспечивается вторым оплодотворением.
Второй спермий пыльцевой трубки, попадая в зародышевый мешок, сливается с диплоидным ядром центральной клетки зародышевого мешка. При этом образуется клетка с тройным набором хромосом: два одинаковых набора хромосом материнского организма и один набор отцовского (рис. 26, 3).
Слияние одного спермия с яйцеклеткой, а другого с ядром-центральной клетки называют двойным оплодотворением. Это открытие сделано в 1898 г. нашим соотечественником С. Г. Навашиным.
Таков в самых общих чертах процесс оплодотворения у животных и растений. Однако он подвержен приспособительным изменениям в зависимости от особенностей строения половых клеток и биологии размножения, свойственных каждому виду животных и растений.
До сих пор рассматривался процесс оплодотворения, связанный с поведением ядер, но, очевидно, этим суть оплодотворения не исчерпывается: оно представляет собой сложный физиологический и биохимический процесс.
Моноспермия, полиспермия, избирательность и селективное оплодотворение. Несмотря на огромное количество сперматозоидов и пыльцевых зерен, приходящихся на одну яйцеклетку животного и растения, оплодотворение осуществляется, как правило, лишь при участии одного сперматозоида и одного пыльцевого зерна. Такой тип оплодотворения называют моноспермным
56
оплодотворением. Он характерен для большинства животных и растений.
Моноспермное оплодотворение контролируется рядом механизмов. Одним из них является блокирование яйца после проникновения в него одного сперматозоида.
Блокирование яйцеклеток у некоторых животных протекает в течение минуты и обеспечивается образованием оболочки оплодотворения и возникновением перивителлинового пространства.
Физиологическая сущность блокирования яйца после соприкосновения головки сперматозоида с поверхностью яйцеклетки не выяснена.
У растений также после проникновения в зародышевый мешок одной пыльцевой трубки развиваются процессы, препятствующие проникновению других.
Однако у ряда животных в цитоплазму яйцеклетки проникает несколько сперматозоидов. Явление проникновения в цитоплазму яйцеклетки нескольких сперматозоидов называется полиспермией.
Полиспермия широко распространена у беспозвоночных: моллюсков, иглокожих, насекомых; встречается она и у позвоночных: рыб (закономерно у акуловых), амфибий, рептилий и птиц. У млекопитающих в норме полиспермия встречается редко (1—2%); исключением являются однопроходные, у которых это явление, возможно, является правилом.
У растений также наблюдаются случаи полиспермии, когда в зародышевый мешок проникает несколько пыльцевых трубок. Полиспермия отмечена у свеклы, хлопчатника, гречихи, табака и других растений.
Однако, несмотря на проникновение в яйцеклетку нескольких сперматозоидов в случае полиспермии, с женским пронук-леусом соединяется только один мужской пронуклеус. Остальные элиминируются. Полиспермная кариогамия, т. е.
слияние нескольких пронуклеусов, в норме не обнаружена.
Описаны случаи, когда у растений дополнительные спермин сливаются не с ядром яйцеклетки, а с другими ядрами зародышевого мешка (синергидами или антиподами), тогда из одного зародышевого мешка развивается несколько зародышей (полиэмбриония).
Проникновение в цитоплазму яйцеклетки нескольких спермиев и слияние с ее ядром только одного из них давно наводили на мысль, что данный процесс не является чисто механическим. Допускается возможность избирательности в процессе кариогамии, т. е. способность женского пронуклеуса сливаться с определенным мужским пронуклеусом.
Яйцеклетки могут преимущественно оплодотворяться сперматозоидами с определенными свойствами так же за счет конкуренции последних. Такое явление называется селективным оплодотворением. Оно приводит к ограничению свободного скрещивания (панмиксии) и является одним из приспособительных механизмов изоляции в эволюции растений и животных.
57
Биология
У Покрытосеменных растений спорогенез, гаметогенез и оплодотворениепредставляют собой непрерывную последовательность репродуктивных процессов, происходящих в цветках. В типичных цветках репродуктивные органы представлены гинецеем (совокупностью плодолистиков, образующих женскую репродуктивную сферу) и андроцеем (совокупностью тычинок, образующих мужскую репродуктивную сферу).
Мегаспорогенез, образование зародышевого мешка и яйцеклетки
Мегаспорогенез протекает в женской репродуктивной сфере – в гинецее. Морфологически гинецей представлен пестиком (или несколькими пестиками). В состав отдельного пестика входят: завязь, столбик и рыльце. Внутри завязи находятся семязачатки (один или несколько).
Внутреннее содержимое семязачатка называется нуцеллус; снаружи семязачаток окружен покровами (одиночным или двойным интегументом). В нуцеллусе семязачатка (2n) содержится одна археспориальная клетка, способная делиться путем мейоза (у некоторых растений археспорий многоклеточный).
В результате мейоза из археспориальной клетки (материнской клетки мегаспор) образуется четыре мегаспоры. Вскоре три из них отмирают, а одна увеличивается в размерах и трижды делится путем митозов. В результате образуется восьмиядерный зародышевый мешок (женский гаметофит).
Три ядра вместе с прилегающей цитоплазмой образуют клетки-антиподы, два ядра – одно центральное диплоидное ядро; два ядра – две клетки-синергиды; одно ядро становится ядром яйцеклетки.
Микроспорогенез и образование пыльцевых зерен
Микроспорогенез протекает в мужской репродуктивной сфере – в андроцее. Каждый элемент андроцея – тычинка – состоит из тычиночной нити и пыльника. Внутри пыльника имеются пыльцевые гнезда, содержащие археспориальную ткань.
В археспориальной ткани пыльников из каждой диплоидной материнской клетки микроспор в результате мейоза образуется четыре микроспоры.
Каждая микроспора делится путем митоза и образуется двухклеточное пыльцевое зерно: одна клетка называется генеративной (в дальнейшем на ее основе образуются спермии), вторая – вегетативной (это остаток вегетативных клеток мужского гаметофита). Зрелое пыльцевое зерно покрыто двойной оболочкой: экзиной (внешней) и интиной (внутренней).
Опыление
Опыление – это процесс переноса пыльцы из пыльников на рыльце пестика. Существует два основных типа опыления: самоопыление и перекрестное опыление. При самоопылении происходит перенос пыльцы в пределах одного цветка, а при перекрестном опылении пыльца из пыльников одного цветка переносится на рыльце пестика другого цветка.
Для более точной характеристики опыления с точки зрения генетики используются специальные термины: клейстогамия (самоопыление в нераскрывшемся цветке), автогамия (самоопыление в раскрывшемся цветке), гейтоногамия (перенос пыльцы из пыльника одного цветка на рыльце пестика другого цветка в пределах одного растения), аллогамия (перенос пыльцы из пыльника цветка одного растения на рыльце пестика цветка другого растения). Таким образом, лишь аллогамия является собственно перекрестным опылением, а остальные типы опыления тождественны самоопылению.
Образование пыльцевой трубки и спермиев
В результате опыления пыльцевое зерно попадает на рыльце пестика и прорастает в пыльцевую трубку (мужской гаметофит). В пыльцевой трубке ядро генеративной клетки делится путем митоза, образуя два генеративных ядра. (У ряда растений деление генеративной клетки происходит еще в пыльниках). Каждое генеративное ядро с прилежащим слоем цитоплазмы называется спермием.
Двойное оплодотворение, образование зародыша и семени
Пыльцевая трубка внедряется в ткани столбика и растет в направлении завязи, при этом на вершине пыльцевой трубки находится вегетативное ядро, а за ним движутся оба спермия. Затем пыльцевая трубка проникает в семязачаток через пыльцевход – микропиле (у некоторых растений существует халазогамия – проникновение пыльцевой трубки в семязачаток через его ножку – халазу).
Далее пыльцевая трубка лопается, ее содержимое изливается в зародышевый мешок, и происходит двойное оплодотворение. Один спермий сливается с ядром яйцеклетки, и образуется диплоидная зигота. Другой спермий сливается с центральным диплоидным ядром зародышевого мешка, и образуется триплоидное ядро эндосперма.
Все остальные ядра зародышевого мешка и вегетативное ядро пыльцевого зерна – разрушаются.
Из диплоидной зиготы развивается зародыш, а из триплоидного центрального ядра – эндосперм. Происходит формирование семени, включающего зародыш, эндосперм и семенную кожуру, которая образуется из покровов семязачатка.
Спорогенез. Гаметогенез
Бесполое
размножениерастений
и грибов
осуществляется спорами, то есть
специализированными клетками, служащими
для размножения и расселения (рис.
42).
Рис.
42.
Споры
некоторых агариковых грибов.
Количество
спор, образуемых одним растительным
или грибным организмом, может быть
весьма
велико. Споры не дифференцированы
в половом отношении. У водорослей и
грибов они диплоидны или гаплоидны.
Споры высших растений всегда гаплоидны.
У части водорослей и многих грибов
споры снабжены ундулиподиями (жгутиками)
и
подвижны – такие споры называются
зооспорами.
Споры
наземных растений обычно не имеют
приспособлений для активного движения
и защищены от высыхания
твердой клеточной оболочкой, состоящей
из двух слоев – тонкого, проницаемого
внутреннего слоя (интины) и толстого,
непроницаемого для воды и газов внешнего
слоя (экзины).
Организм,
производящий споры, получил название
спорофита,а
процесс образования спор – спорогенеза.У
растений споры образуются в спорангиях
(если зооспоры – то в зооспорангиях).
Спорангий водорослей (низшие растения)
обычно одноклеточный; содержимое этой
единственной клетки делится, в
результате чего формируются споры,
которые
выходят наружу при вскрывании оболочки
спорангия.
У высших растений спорангий
– многоклеточный орган, имеющий
одно- или многослойную стенку. Внутри
многоклеточного спорангия формируется
образовательная ткань – археспорий
(от греческого «архе» – начало), из
которой
в дальнейшем образуются споры. Из
клеток археспория споры возникают в
результате
мейоза.
На гаплоидном организме
(некоторые грибы) споры образуются
в результате митотического деления.
При прорастании такие споры воспроизводят
новую особь, идентичную материнской.
На диплоидном организме формированию
спор предшествует мейоз.
Из
спор, образовавшихся в результате
мейоза, при прорастании возникает
гаплоидный организм, который будет
размножаться половым способом (за счет
образования гамет) – гаметофит
(заросток),не
идентичный диплоидному материнскому
спорофиту.
У
низших растений, мхов и даже у многих
современных высших сосудистых споровых
растений (хвощей, плаунов и папоротников)
все споры одинаковы по размерам и
физиологическим особенностям. Это
равноспоровые
организмы.
У многих растений (некоторые
плауны, папоротники, все голосеменные
и цветковые)
на одной и той же особи или на разных
особях одного вида образуются споры
двух типов, различные по размерам и
физиологическим особенностям. Такие
растения называются разноспоровыми
(некоторые плауны и папоротники, все
голосеменные и покрытосеменные).
Относительно
более мелкие споры (микроспоры)
(рис.
43)
образуются в
микроспорангии, а более крупные
(мегаспоры)
–
в мегаспорангии. Микроспоры, прорастая,
дают начало однополому мужскому
гаметофиту (мужской особи), на котором
возникают мужские половые органы –
антеридии.
Мегаспоры
при прорастании образуют женский
гаметофит (женскую особь), несущий
женские половые органы – архегонии.Разноспоровость
эволюционно возникла у высших растений
(некоторые плауновидные и папоротники,
все голосеменные и покрытосеменные).
Рис. 43.
Микроспоры
цветковых растений
. А – магнолия, Б – сусак, В – джузгун, Г – пион, Д – лютик, Е – истод.
Половое
размножение
– это
такой тип размножения, при котором новые
особи образуются в результате полового
процесса.
Для полового процесса
необходимы, как правило, две родительские
особи, продуцирующие два физиологически
различных типа половых клеток (гамет)с
перекомбинированными родительскими
хромосомами, которые сливаются и
формируют зиготу.
Из зиготы в последующем развивается
новая дочерняя особь
Организмы,
на которых формируются гаметы, называются
гаметофитами, а процесс образования
гамет – гаметогенезом.У
большинства растений и грибов гаметы
возникают в особых органах – гаметангиях.Гаметы
всегда гаплоидны.
Гаметофиты равноспоровых
растений обычно обоеполы и формируют
как мужские, так и женские гаметангии.
У разноспоровых растений обычно из
микроспор вырастают гаметофиты с
мужскими гаметангиями, а из мегаспор –
гаметофиты с женскими гаметангиями.
В связи
с тем, что разные растения способны
вырабатывать различные по величине и
подвижности типы гамет, различают
несколько типов полового процесса. В
простейшем случае у некоторых одноклеточных
водорослей и грибов, лишенных твердой
оболочки, сливаются целые одноклеточные
организмы. Такой половой процесс
называется хологамией.
Если
в половом процессе участвуют
специализированные половые клетки –
гаметы, то такой процесс называется
гаметогамией (рис.
46).
Гаметы, образующиеся в гаметангиях,
могут быть морфологически однотипны,
тогда их называют изогаметами.
Попарное
их слияние основано лишь на физиологическом
различии, а половой процесс такого типа
называют изогамным. Изогамия
встречается
у некоторых водорослей и у очень немногих
грибов.
Если подвижные гаметы различаются
по величине (одна превосходит другую
в несколько раз), то слияние таких гамет
обусловливает гетерогамный
половой
процесс. У некоторых грибоподобных
организмов (оомицеты), водорослей и у
всех высших растений половой процесс
оогамный.
Женская
гамета (яйцеклетка) при оогамной форме
полового процесса неподвижна, лишена
жгутиков (ундулиподиев), имеет крупные
размеры и большой запас питательных
веществ.
Мужская гамета (сперматозоид)
– маленькая, подвижная, со жгутиками
(ундулиподиями), либо без жгутиков (тогда
она называется спермий); состоит из
крупного ядра и очень небольшого
количества цитоплазмы. Неподвижная
яйцклетка образуется либо в гаметангии,
называемом оогонием (у водорослей и
некоторых грибов), либо в архегонии
(высшие растения, исключая цветковые).
Оогоний
(от
греческого «оон» – яйцо,
«гоне» – рождение) обычно состоит из
одной клетки, реже (у харовых водорослей)
многоклеточный. Архегоний(от
греческого
«архе» – начало) – женский половой
орган высших архегониальных растений
– мхов, плаунов, хвощей, папоротников
и голосеменных.
Это всегда многоклеточное
образование, состоящее из
расширенного брюшка, в котором помещается
яйцеклетка, и удлиненной шейки.
Над яйцеклеткой находится брюшная
канальцевая
клетка. Наружные клетки архегония
стерильны и образуют однослойную стенку.
По каналу шейки, заполненному
слизью, сперматозоиды проникают в
брюшко и один из них сливается с
яйцеклеткой.
Сперматозоиды
у споровых растений образуются
в гаметангиях, называемых антеридиями.
Антеридий(от
греческого «антерос»
– цветущий) – это одноклеточный (у
большинства низших растений) или
многоклеточный (у высших споровых
растений)
мужской половой орган, имеющий
у разных систематических групп различную
(округлую или овальную) форму.
Сперматозоиды,
созревшие в антеридиях,
могут достичь яйцеклетки только в
присутствии капельно-жидкой воды.
Наличие
ее – обязательное условие для
осуществления
процесса оплодотворения у
всех групп растительных организмов,
исключая
семенные растения.
У цветковых растений
произошла полная редукция гаметангиев,
однако половые клетки (спермии и
яйцеклетки) образуются и участвуют в
половом процессе.
Помимо
типичного полового процесса,
в котором обязательно участвуют две
гаметы,
существует особый тип полового процесса,
при котором зародыш развивается
из неоплодотворенной яйцеклетки.
Это
явление у растений чаще всего известно
под названием апомиксиса(от
греческого
«апо» – от (без) и «миксис» – смешение).
Апомиксис широко встречается у
многих покрытосеменных растений (часть
розоцветных, сложноцветных), особенно
у ряда культурных растений (свекла,
хлопчатник, лен, табак).
Рис.
44.
Различные типы полового размножения
у растений, водорослей и грибов (схема).
А
–
изогамия (характерна
для некоторых водорослей); Б
–
гетерогамия (некоторые
водоросли); В
–
оогамия (некоторые водоросли,
все высшие растения); Г – конъюгация
(некоторые
водоросли).
Половая
и бесполая формы размножения
имеют определенные биологические
преимущества. При
половом размножении достигается
комбинация наследственного материала
родительских
форм.
Образующаяся при половом
воспроизведении особь генетически
нетождественна ни одному из своих
родителей.
Половой процесс обеспечивает
генетическую изменчивость организмов
из поколения в поколение, а половое
размножение дает виду преимущества,
реализующиеся при естественном
отборе.
При бесполом размножении
наследственные особенности передаются
без изменений и могут легко закрепляться
в ряду поколений. Однако чаще
всего у растений осуществляется
чередование
полового и бесполого поколений.
При чередовании поколений гаметофит
закономерно сменяется спорофитом,
который
затем вновь сменяется следующим в череде
поколений гаметофитом.
Гаметофит
и спорофит могут быть одинаковыми
как морфологически, так и
по продолжительности жизни (изоморфное
чередование поколений) (у
многих водорослей) или
резко
различными (гетероморфное чередование)
(у некоторых водорослей и всех высших
растений).
Таким
образом, у разных групп водорослей
встречаются обе формы
чередования поколений. Для высших
растений характерно только гетероморфное
чередование. Оба поколения развиваются
либо независимо как самостоятельные
особи, либо одно поколение «поселяется»
на другом. Так, у мхов спорофит
развивается на зеленом гаметофите.
У семенных растений крайне редуцированный
бесхлорофилльный женский гаметофит
не покидает спорофит.
У
всех высших растений, кроме мхов,
преобладает спорофит, а гаметофит развит
слабее и недолговечен.
Чередование поколений связано со сменой
ядерных фаз – гаплоидной (n)
и диплоидной
(2n).
Эта смена обеспечивается благодаря
мейозу,
протекающему у всех высших растений
при
спорогенезе,
и половому процессу.
Диплоидный спорофит высших растений всегда
производит гаплоидные мейоспоры. Из
них вырастает гаплоидный гаметофит,
продуцирующий гаплоидные митогаметы
(половые клетки, образованные в процессе
митоза). При слиянии гамет восстанавливается
диплоидное
число хромосом
в зиготе, из которой вырастает
диплоидный спорофит.
Задание 1
1. Заполните таблицу
предоставив общую характеристику
бесполого и полового размножения.
Таблица 3 – Общая
характеристика бесполого и полового
размножения
| Показатель | Способ размножения |
| бесполое | половое |
| Клеточные источники наследственнойинформации | |
| Родители | |
| Потомство | |
| Главный клеточный механизм | |
| Эволюционное значение |
Задание 2
1. Спорообразование
у плесневого гриба. Нанесите пипеткой
каплю воды на предметное стекло,
возьмите пинцетом несколько нитей мицелия плесневого гриба, положите их в каплю воды на предметное стекло и накройте покровным стеклом.
Рассмотрите препарат под малом и большом увеличением микроскопа. В поле зрения видны ветвящиеся полупрозрачные
и тонкие нити – гифы. Найдите среди гиф спорангии – круглые черные коробочки на длинной ножке (спорангиецосце), заполненные мелкими спорами округлой формы.
Некоторые из спорангиев могут оказываться разорванными, и в таких случаях вокруг них видны высыпавшиеся
мелкие округлые клетки – споры. Внутри спорангия обнаруживается колонка – утолщенный вырост спорангиеносца; форма и размеры колонки постоянны у
определенных видов грибов.
Зарисуйте
мицелий плесневого гриба, на рисунке
обозначьте: мицелий, гифы, спорангий,
споры.