Научная сенсация: в МГУ разгадали механизм упаковки молекулы ДНК

Исследователи лаборатории биологического факультета МГУ стали авторами научной сенсации. Им удалось понять, как нить ДНК длиной в два метра помещается в ядро диаметром не более одной сотой доли миллиметра. При дополнительном анализе это открытие позволит выявить способы лечения многих серьезных заболеваний, в том числе и рака.
В лаборатории биологического факультета МГУ сделано открытие, о котором сейчас говорит весь научный мир. Российским ученым удалось понятьмеханизмы упаковки генома внутри клетки.
До сих пор не вполне ясно, почему клетки, например, кожи или сетчатки глаза, отличаются друг от друга, хотя все содержат одинаковый геном. Оказывается, это может зависеть и от того, как носитель генетической информации — молекула ДНК — уложена в пространстве.
“ДНК, в которой закодированы все наши гены, очень длинная — ее длина составляет порядка двух метров, а размер ядра, в котором она находится, составляет порядка 10 микрон. Соответственно, она должна быть очень компактно уложена, чтобы просто поместиться в это ядро.
Но проблема — существенно более сложная, потому, что она должна не просто там быть уложена, а быть уложена таким образом, чтобы те ее части, которые сейчас, на данный текущий момент, востребованы, или в данных клетках востребованы, были доступны для тех машин, которые, скажем, считывают информацию с этой ДНК”, — рассказывает заведующий кафедрой молекулярной биологии МГУ Сергей Разин.
Чтобы длинная молекула ДНК не запуталась, она накручивается на особую структуру из белков в форме бочонков. Но геном — это не крепкий клубок ниток. На каком-то участке ДНК, оказывается, намотана плотно, на другом — нет. Исследователи считают, что плотность упаковки напрямую связана с работой генов.
“В этом проекте мы впервые в нашей лаборатории и, насколько мне известно, впервые в России использовали метод, который позволяет получить карту трехмерной организации всего генома.
Если гены работают сильно, то этот участок расправляется, если эти гены молчат в другой клеточной линии, то этот участок оказывается “схлопнутым” в более плотную конфигурацию”, — говорит старший научный сотрудник кафедры молекулярной биологии МГУ Сергей Ульянов.
Слипаются и разлипаются участки ДНК под действием особых ферментов там, где это нужно в данный момент для строительства новой клетки.
Подтвердить полученные данные помогло моделирование биологического процесса на суперкомпьютере “Ломоносов”. За секунду каждый его процессор совершает тысячи миллионов операций.
Чтобы рассчитать модель работы ДНК, потребовалось 200 таких процессоров и три дня работы.
“Приятно, когда статистическая физика описывает такие сложные и, казалось бы, плохо поддающиеся статистической физике объекты, как живая природа, которая состоит сплошь из исключений, а не из правил.
Эта структура очень живая — вот ее моментальная фотография в какой-то один момент времени.
Если вы посмотрите на нее в другой момент времени, она будет выглядеть очень мало похожим образом”, — объясняет научный сотрудник Физического факультета МГУ Александр Чертович.
Структура, которая готова в любой момент поменяться, в частности, объясняет, почему живой организм так быстро может отреагировать на изменение окружающей среды.
В будущем новое открытие поможет ученым и понять причины возникновения многих заболеваний, в том числе и рака, который как раз является следствием нарушения генетического кода.
Но для получения таблеток, корректирующих работу генов, нужно провести еще немало научных опытов.
Учёные пришли к выводу, что укладка в особое состояние под названием “фрактальная глобула” позволяет всей этой генетической машинерии клетки работать с максимальным быстродействием.
Фрактальная глобула – понятие математическое. Но его можно объяснить на примере верёвки или лески. Если вы уроните на пол длинную рыболовную леску (например, от спиннинга), она тут же свернется в такой невообразимо “подлый” клубок, что вам придётся либо распутывать его часами, либо бежать в магазин за новой катушкой. Это обычная, так называемая равновесная, глобула.
Фрактальная глобула – структура в этом смысле намного более “вежливая”. Применительно к леске это комок, в котором леска ни разу не завязалась в узел, она просто свернулась множество раз, так, чтобы ни одна петля не обвилась вокруг другой. Такая структура представляет собой множество свободных петель разного размера – потяни её за два конца, и она легко распутается.
Российские физики Александр Гросберг, Сергей Нечаев и Евгений Шахнович, впервые спрогнозировавшие существование её ещё в 1988-м году, назвали такую глобулу “складчатой”. Всё потому, что она чем-то похожа из-за своей укладки на современную лапшу быстрого приготовления в виде брикетов (“доширак”),
В последние годы её чаще называют фрактальной – и звучит научнее, да и полней отражает свойства такой глобулы, поскольку, как и во всех фракталах, её структура (в данном случае форма мелких и крупных петель) повторяется на малых и больших масштабах.
Долгое время это предсказание оставалось невостребованным. Но результаты исследований последних лет указывают, что хромосомы (нити ДНК) складываются в ядре именно в такую конфигурацию – во фрактальную глобулу.
Сегодня по этому поводу в научном сообществе нет консенсуса.
Но большинство специалистов, работающих в этой области, сильно заинтригованы, и последние 5-7 лет наблюдается целый поток исследований, посвященных геному, свёрнутому во фрактальную глобулу.
Интуитивно это было бы понятно. Двойная спираль ДНК, укреплённая соответствующим набором белков, представляет собой длинную нить, называемую хроматином.
И если этот хроматин представляет собой библиотеку технических руководств по синтезу того или иного белка, нужного организму, то лучше было бы текст этих руководств без нужды не трогать.
Соответственно, организму нужно избегать ненужных перекрещиваний одного гена с другим и складывать хроматиновую нитку так, чтобы ни в одном месте части этой нитки между собой не завязывались узлом.
Поэтому, как бы эта нитка хроматина в ядре ни складывалась, она не должна повторять судьбу нечаянно упавшей на пол рыболовной лески, то есть быть не простой глобулой, а фрактальной.
Вдобавок нитка во фрактальной глобуле, не имеющая узлов, по идее должна иметь более высокую свободу перемещений, что для ДНК немаловажно.
Для того чтобы ДНК нормально функционировала, необходимо, чтобы её отдельные части в нужный момент встречались между собой, “включая” сигнал к считыванию и указывая всей системе место, откуда это считывание следует начинать, причем всё это должно происходить достаточно быстро (в том числе и во время починки главного носителя информации).
Учёным из МГУ удалось придумать и посмотреть на поведение модельной (более короткой чем реальная ДНК) цепи, свёрнутой во фрактальную глобулу.
У их предшественников смоделировать ситуацию с длинной цепочкой, состоящей из четверти миллиона звеньев, не получалось.
По признанию исследователей, моделирование длинных цепочек (а именно они позволяют получить сколько-нибудь значимые результаты) затрудняется тем, что они очень долго приходят в равновесное состояние.
“Мы сумели оценить тепловую динамику, свойственную этому виду укладки. Проведённое нами компьютерное моделирование хорошо подтвердило теоретический результат”, – отметил Михаил Тамм, сотрудник кафедры физики полимеров и кристаллов физического факультета МГУ, один из авторов исследования.
Удачно разрешив эту проблему за счёт грамотно выстроенной программы и большого компьютерного времени на суперкомпьютере МГУ “Ломоносов”, исследователи смогли оценить динамику теплового движения (изменение в движении, происходящем за счёт температуры) в моделируемой фрактальной глобуле.
Оказалось, что звенья хроматиновой цепочки движутся во фрактальной глобуле быстрее, чем в обычной (“запутанной”). Что в числе прочего могло повлиять на выбор природы (в ходе эволюционного развития) фрактальной глобулы в качестве лучшего способа укладки ДНК в ядре.
Исследователи надеются, что их работа позволит лучше понять, как именно функционирует вся машинерия, связанная с хранением и считыванием информации в ДНК.
“С точки зрения динамики нам бы хотелось разобраться с тем, какие там встроенные характерные времена, какие процессы могут происходить просто за счёт теплового движения, а что неизбежно требует привлечения активных элементов, ускоряющих работу ДНК”, – резюмировал Тамм.
http://www.vesti.ru/doc.html?id=2694727
Похожее
comments
Источник: https://novostik.com/nauchnaya-sensaciya-v-mgu-razgadali-mexanizm-upakovki-molekuly-dnk/
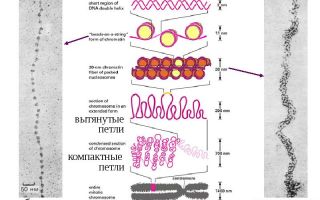
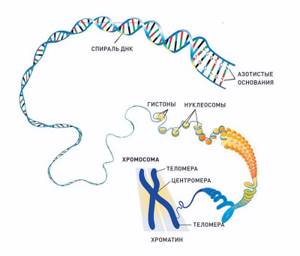
Уровни упаковки ДНК. Строение хромосом
Упаковка ДНК включает несколько уровней (рис. 11).
Первый уровень заключается в образовании нуклеосомной нити. Нуклеосома состоит из октамера белков-гистонов, обвитого 2 витками ДНК. Октамер (греч.
octo – восемь, meros – часть) состоит из 8 глобулярных белков – гистонов (парные Н2А, Н2В, Н3, Н4). Нуклеосомы разделены короткими участками линкерной ДНК. С октамером и частично с участком линкера связывается гистон Н1.
Цепь нуклеосом называется нуклеосомной нитью. Она имеет диаметр ~10 нм. ДНК укорачивается в 6 – 7 раз.
Рис. 11. Упаковка ДНК. Строение метафазной хромосомы
Второй уровень упаковки – образование хроматиновой фибриллы. При этом нуклеосомная нить закручивается в спираль диаметром 30 нм. ДНК укорачивается в 40 раз. В дальнейшей упаковке хроматина ведущую роль играют негистоновые белки.
Третий уровень упаковки ДНК – хромомерный. Хроматиновые фибриллы образуют петли – хромомеры, диаметром около 100-150 нм. ДНК укорачивается в 600 раз. Нитчатая хроматиновая структура со средней толщиной 100-200 нм получила название хромонемы. Хромонему можно наблюдать на стадии начальной конденсации хромосом в профазе митоза и при деконденсации хромосом в телофазе.
Четвертый уровень – хромосомный. Заключается в более плотной упаковке петель и образовании «конденсированных участков хромосом» диаметром около 700 нм. Максимальной степени конденсации хромосомы достигают в метафазе митоза/меойза.
Строение хромосом (рис. 11)
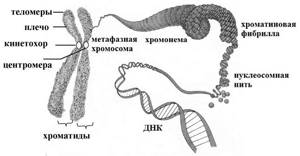
Хромосомы на стадии метафазы образованы двумя хроматидами, объединенными в области первичной перетяжки – центромеры. Каждая хроматида представляет собой дочернюю ДНК. В зонах центромер расположены белковые структуры – кинетохоры, они являются местом прикрепления нитей веретена деления. Участки, на которые центромера делит хромосому, называются плечи, концы хромосомы – это теломеры.
Некоторые хромосомы имеют вторичные перетяжки (ядрышковые организаторы), они содержат гены, кодирующие рРНК. Часть хромосомы, отделенная вторичной перетяжкой, называется спутник или саттелит.
По положению центромеры хромосомы делятся на 3 вида (рис. 12):
1) метацентрические – хромосомы, которые имеют равные плечи;
2) субметацентрические – хромосомы с неравными плечами;
3) акроцентрические – хромосомы, у которых одно плечо практически отсутствует.
Рис. 12. Форма хромосом:
А – метацентрическая хромосома, Б – субметацентрическая хромосома,
В – акроцентрическая хромосома
Источник: http://www.compendium.su/biology/total/9.html
Как омолодить организм на клеточном уровне, подскажет
Ученые выяснили детали механизма клеточного старения, запускающегося по мере деления клеток и вызывающего изменения в структуре упаковки ДНК, что может быть использовано для разработки новых методов омоложения стареющего организма, сообщает журнал Nature Structural and Molecular Biology.
Старение клетки, в отличие от старения людей или животных, заключается не в постепенном угасании, дряхлении и утрате способности выполнять какие либо функции, а в том, способна ли клетка совершить очередной акт деления. С течением времени скорость деления клеток замедляется до тех пор, пока этот процесс не прекращается совсем.
Именно прекращение подпитки организма новыми клетками в самых разных типах тканей и приводит к его старению в целом.
До сих пор, однако, ученым неизвестны детальные причины замедления деления клеток и методы обращения этого процесса вспять или хотя бы его замедления.
Считается, что за старение каким-то образом отвечают теломеры – концевые участки хромосом, укорачивающиеся каждый раз при акте деления клетки .
“До нашего исследования мы знали, что теломеры становятся короче по мере того, как клетка делится, и когда они становятся слишком короткими, клетки либо перестают делиться, либо вовсе отмирают.
Таким образом, что-то должно, по нашему мнению, передавать сигнал от теломер ко всему клеточному ядру, который бы запускал какие-то изменения в работе ДНК в целом”, – сказал соавтор публикации Ян Карлсендер (Jan Karlseder) из Солковского института биологических исследований, слова которого приводит пресс-служба этой организации.
Карсендер и его коллеги провели сравнения в количестве особых белков гистонов между старыми и молодыми клетками, выращиваемыми в искусственных условиях. Гистоны образуют комплексное соединение с молекулами ДНК в ядре, помогают им принять определенную форму трехмерную структуру упаковки, образуя так называемый хроматин – вещество на основе ДНК, из которого состоят хромосомы.
Очень быстро команда Карсендера обнаружила, что старые клетки, претерпевшие около 75 актов деления, содержат в своем ядре заметно меньше гистонов по сравнению с молодыми, делившимися всего около 30 раз, передает РИА Новости.
“Эти белки выполняют вспомогательную функцию во всех участках ДНК, а потому ее нарушение из-за недостатка гистонов неизбежно сказывается на работе всего генома”, – сказал соавтор статьи Роди О'Салливан (Roddy O'Sullivan).
Проведя более тщательный анализ, ученые обнаружили, что негативные изменения в количестве и разнообразии гистонов наблюдались у клеток на всех стадиях их жизни в зависимости от количества осуществленных процессов деления.
Эта работа демонстрирует, что старение, вопреки широко распространенному убеждению, будто возрастные болезни наступают из-за накопления повреждений в самой ДНК, является намного более сложным процессом. Процесс старения может ускоряться за счет укорочения теломер, искажающих взаимодействие генов с гистонами и таким образом нарушающих их работу.
Также ученые показали, что генетическое вмешательство в клетку с тем, чтобы вернуть ей способность производить дополнительные количества гистонов, позволило заметно повысить количество и разнообразие этих белков даже в старых клетках. Именно так, по мнению авторов, можно добиться продления жизни клеток.
“Если мы будем просто искусственным путем удлинять теломеры, мы можем перевести их в так называемое “бессмертное” состояние, свойственное раковым клеткам”, – пояснил Карлсендер.
Источник: http://sbio.info/news/newsbioteh/kak_omolodit_organizm_na
Молекулярные биологи научились исправлять ошибки упаковки ДНК
Этот подход может стать революционным в лечении различных генетических заболеваний у людей.
Молекулярные биологи из ведущих американских университетов – Медицинской школы Бейлорского университета, Стэндфордского, Гарвардского университетов, Массачусетского технологического института разработали новый метод, с помощью которого можно редактировать геном. Или, как говорят сами авторы, – делать «операции» на уровне генома.
Метод основан на изменении упаковки ДНК в хромосомах.
Длина молекулы ДНК составляет около двух метров, и такая огромная структура помещается в крошечной хромосоме в ядре клетки. Добиться такой компактности удается за счет упаковки. Упаковка происходит так, что спираль ДНК наматывается на комплекс определенных белков, как на катушку. Количество изгибов превышает десять тысяч.
Недавно молекулярным биологам из Медицинской школы Бейлорского университета удалось выяснить, что аномалии упаковки и скручивания ДНК приводят к различным генетическим заболеваниям.
Причем, как показывали исследования, скручивание определяется тем, как основания на концах изгибов объединяются с одним из ДНК-связывающих белков – так называемых «цинковых пальцев».
В данном случае белком – «цинковым пальцем» CTCF.
«Цинковые пальцы» – это группа ДНК-связывающих белков. В их структуре имеется характерный домен, включающий два цистеиновых и один гистидиновый остаток. Эти аминокислоты взаимодействуют с ионом цинка, а расположенная между ними полипептидная цепочка образует петлю в форме пальца. Цинковые пальцы – это регуляторы транскрипции, иными словами, регуляторы работы гена.
В своем нынешнем исследовании ученые совершили большой шаг вперед – им удалось показать, что изменяя последовательность на изгибах ДНК, можно влиять на сам процесс укладки ДНК и формирование изгибов.
«Это означает, что теоретически возможно устранять ошибки в укладке ДНК, при этом никак не повреждая соседние участки», – говорит ведущий автор исследования доктор Эрез Эйден (Erez Lieberman Aiden).
Для редактирования укладки ДНК авторы использовали так называемую CRISPR-систему. В основе работы этой системы лежит белок Cas9, разрезающий двуцепочечную цепь ДНК клетки, чтобы туда можно было вставить нужный отрезок. С помощью РНК-гидов, коротких направляющих последовательностей РНК, генетики научились показывать Cas9 место, где необходимо выполнить разрез.
«Использование этой системы позволило нам работать, как будто «генетическим скальпелем» и добавлять или удалять различные небольшие участки генетического кода. Эти манипуляции дают возможность изменять укладку ДНК, причем, точно добиваясь той укладки, которая нам нужна», – говорит один из авторов исследования аспирант Сухас Рао (Suhas Rao).
«Белок CTCF работает, как код, который и определяет скручивание ДНК. Теперь, когда нам удалось разгадать этот код, мы можем контролировать и весь процесс укладки ДНК», – говорит один из авторов исследования Эрик Ландер (Eric Lander).
Более подробно о результатах этого исследования можно прочитать в свежем выпуске журнала PNAS.
Источник: https://www.infox.ru/news/84/science/medicine/153035-molekularnye-biologi-naucilis-ispravlat-osibki-upakovki-dnk
упаковка [ДНК] – это… Что такое упаковка [ДНК]?
- упаковка [ДНК] — Совокупность процессов спирализации и самоукладки двухцепочечной молекулы ДНК, ведущих к резкому сокращению ее абсолютной длины; эффективность У. оценивается по индексу упаковки. [Арефьев В.А., Лисовенко Л.А. Англо русский толковый словарь… … Справочник технического переводчика
- Гибридизация ДНК — У этого термина существуют и другие значения, см. гибридизация. Гибридизация ДНК, гибридизация нуклеиновых кислот соединение in vitro комплементарных одноцепочечных нуклеиновых кислот в одну молекулу. При полной комплементарности… … Википедия
- ПОЛИМЕРЫ БИОЛОГИЧЕСКИЕ — (биополимеры) природные макромолекулы, играющие осн. роль в биол. процессах. К П. б. относятся белки, нуклеиновые кислоты (НК) и полисахариды. П. б. образуют структурную основу всех живых организмов; все процессы в клетке связаны с… … Физическая энциклопедия
- Деацетилазы гистонов (HDAC) — Деацетилазы гистонов (Histone deacetylases, HDACs), К. Ф. 3.5.1 ферменты катализирующие удаление ацетильной группы ε N ацетил лизина гистонов, внесенные ферментами гистонацетилазами (histone acetylases, HATs) в остатки K3 и K14 гистона Н3 и … Википедия
- Деацетилаза — Деацетилазы гистонов (Histone deacetylases, HDACs), К. Ф. 3.5.1 ферменты катализирующие удаление ацетильной группы ε N ацетил лизина гистонов, внесенные ферментами гистонацетилазами (histone acetylases, HATs) в остатки K3 и K14 гистона Н3 и K5,… … Википедия
- Деацетилазу гистонов (HDAC) — Деацетилазы гистонов (Histone deacetylases, HDACs), К. Ф. 3.5.1 ферменты катализирующие удаление ацетильной группы ε N ацетил лизина гистонов, внесенные ферментами гистонацетилазами (histone acetylases, HATs) в остатки K3 и K14 гистона Н3 и K5,… … Википедия
- Деацетилазы гистонов — Каталитический домен HDAC4, связанный с ингибитором (показан зелёным цветом). Деацетилазы гистонов (Histone deacetylases, HDACs), К. Ф. 3.5.1 ферменты катализирующие удаление ацетильной группы ε N ацетил лизина гистонов, внесенные… … Википедия
- Протисты — Научная классификац … Википедия
- packing — packing. См. упаковка [ДНК]. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
- Полимер — (Polymer) Определение полимера, виды полимеризации, синтетические полимеры Информация об определении полимера, виды полимеризации, синтетические полимеры Содержание Содержание Определение Историческая справка Наука о Полимеризация Виды… … Энциклопедия инвестора
Источник: https://dic.academic.ru/dic.nsf/genetics/9632/%D1%83%D0%BF%D0%B0%D0%BA%D0%BE%D0%B2%D0%BA%D0%B0
Упаковка ДНК в хромосомах
В клетках или вирусах ДНК, по-видимому, никогда не находится в свободной, вытянутой форме. Она связана с низкомолекулярными катионами — ионами двухвалентных металлов либо с ди- и полиаминами или белками, а возможно, с теми и с другими.
Взаимодействие осуществляется с помощью электростатических сил — отрицательно заряженные фосфатные группы частично нейтрализуются положительно заряженными ионами металлов и полиаминами или основными аминокислотными остатками белков.
В результате таких взаимодействий происходит конденсация ДНК с уменьшением объема, занимаемого молекулой, иногда в тысячу раз. Кольцевая ДНК Е.
coli длиной 1,4 мм заключена в клетку, имеющую форму палочки диаметром 1 мкм и длиной 2 мкм; у эукариотических клеток ядерная ДНК длиной почти 2 м в стадии интерфазы заключена в ядре диаметром менее 10 мкм. Ядерная ДНК в клетках, находящихся в стадии митоза, конденсирована еще больше и в световом микроскопе имеет вид очень компактной структуры.
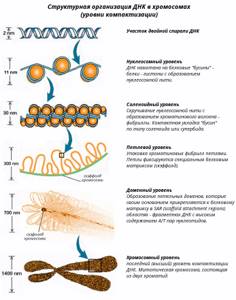
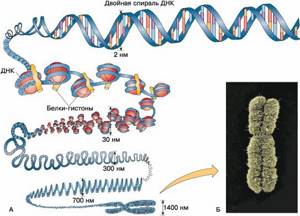
Упаковка ДНК в ядре
В средней эукариотической клетке общая протяженность геномной ДНК составляет около 2 м, диаметр ее ядра всего ~10-20 мкм. При этом совокупность генов, работающих в данной клетке, должна быть доступна для РНК-полимераз и транскрипционных факторов, а вся ДНК в делящихся клетках должна реплицироваться.
Сегодня известно, что упаковка ДНК в ядре эукариотической клетки осуществляется в несколько этапов . Сначала нить ДНК укладывается в нуклеосомы, при этом ее длина уменьшается в шесть-семь раз.
Затем нуклеосомная нить складывается в так называемую 30 нм фибриллу (соленоид или зигзагообразную нить), что обеспечивает дополнительную компактизацию в 40 раз. Далее фибрилла организуется в большие (50 и более тысяч пар нуклеотидов) петли, концы которых закрепляются на белковом скелете ядра (его часто называют ядерным матриксом).
На этом этапе линейные размеры ДНК сокращаются в 700 раз. Существуют и следующие уровни компактизации ДНК, информация о которых в настоящее время весьма скудна и противоречива.
Правильная упаковка ДНК с хромосомными белками осуществляется под наблюдением вспомогательных ферментов. Для корректировки упаковки ферменты-помощники используют энергию АТФ.
Исследователи из Университета Пенсильвании (США) сумели искусственно воспроизвести сворачивание хромосомы, и, как говорят учёные, решающим фактором оказалась энергия — наличие в реакционной смеси молекул АТФ.
Результаты экспериментов опубликованы в журнале Science.
Рис. 1. Уровни упаковки ДНК в ядре эукариотической клетки.
Пока речь шла лишь об упаковке одной протяженной молекулы ДНК. В первом приближении таковой можно считать ДНК одной хромосомы. Однако геном эукариотической клетки разделен на несколько хромосом. Например, в клетках плодовой мушки дрозофилы – имеется четыре пары хромосом (в клетках человека их 46).
Индивидуальные хромосомы можно увидеть под микроскопом только во время митоза. На остальных фазах клеточного цикла они не видны, и ядро клетки представляется относительно гомогенным.
В течение многих лет молекулярных биологов интересовал вопрос, занимают ли отдельные хромосомы ограниченные пространства внутри ядра или же при декомпактизации хромосом ДНК каждой из них распределяется по всему ядру, неизбежно перемешиваясь с ДНК других хромосом. Около 10 лет назад ответ на этот вопрос был найден.
Методы молекулярной гибридизации позволили окрашивать в интерфазном ядре индивидуальные хромосомы .
Оказалось, что они, вопреки общепринятой в то время точке зрения, занимают внутри ядра ограниченные неперекрывающиеся пространства (названные “хромосомными территориями” и располагаются неслучайным образом: хромосомы, богатые генами, локализуются ближе к центру ядра, а бедные генами – ближе к его периферии. В поддержании специфических позиций хромосомных территорий важную роль играет ядерный матрикс.
Хромосомы эукариот
Хромосомы эукариотических клеток состоят в основном из хроматина — комплекса двухцепочечной ДНК и пяти гистоновых белков, обозначаемых H1, Н2А, Н2В, Н3 и Н4.
Гистоны могут быть ацетилированы, метилированы, фосфорилированы, роlу(АDР)-рибозилированы, а гистоны Н2А и Н2В — ковалентно связаны с белком, называемым убиквитином. Какова роль воздействия указанных компонентов на структуру и функции гистонов — до конца не выяснено.
Гистон H1 млекопитающих состоит из примерно 215 аминокислот; размеры других гистонов варьируют от 100 до 135 аминокислот.
Все они содержат необычно большое количество положительно заряженной аминокислоты лизина; Н3 и Н4 отличаются от других тем, что у них достаточно высок уровень положительно заряженной аминокислоты аргинина. Соотношение между Н2А, Н2В, Н3 и Н4, содержащимися в хроматине низших эукариот (дрожжи, плесневые грибы), такое же, как в хроматине млекопитающих.
На электронно-микроскопических фотографиях в зависимости от условий выделения и степени растяжения хроматин выглядит либо как длинное волокно диаметром 10 нм, либо чаще как более вытянутое волокно с утолщениями — «бусинками» диаметром 10 нм, нанизанными по всей длине волокна с определенными интервалами:
Электронные микрофотографии хроматина. А. Волокно хроматина диаметром 10 нм из почечных клеток CV1 обезьяны.
Б. Хроматин из эритроцитов цыпленка, имеющий вид нити с нанизанными на нее бусинками.
Каждая из этих бусинок представляет собой нуклеосомный кор, на который намотан сегмент хромосомной ДНК длиной 145 пар оснований. Кор — это гистоновый октамер, состоящий из гистонов Н2А, Н2В, Н3 и Н4, по две молекулы каждого вида:
Модель нуклеосомного кора, построенная по данным кристаллографического анализа низкого и высокого разрешения.
Сегмент ДНК (145 пар оснований), изображенный в виде трубки, обвивает гистоновый октамер, делая вокруг него 13/4 оборота
Молекула ДНК, обвиваясь 13/4 раза вокруг нуклеосомного кора, образует сверхспираль.
Пятый гистон, H1, не входит в состав нуклеосомного кора и не участвует в процессе наматывания ДНК на гистоновый октамер. Он контактирует с ДНК в тех местах, где двойная спираль входит и выходит из нуклеосомного кора:
Гистон Н1 «сшивает» ДНК в местах, где она начинает и прекращает наматываться на нуклеосомный кор
В такой структуре с одним гистоновым октамером и молекулой гистона H1 ассоциированы 168 пар оснований спиральной ДНК.
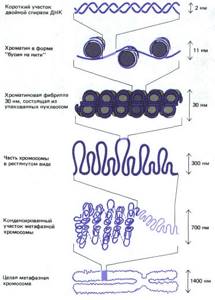
Как мы уже отмечали, на электронно-микроскопических фотографиях хроматин часто обнаруживается в двух альтернативных формах: в форме волокна с четко разделенными нуклеосомами (нуклеосомы имеют вид бусинок, нанизанных на нитку) или в форме волокна диаметром 10 нм, в котором нуклеосомы упакованы бок о бок по всей его длине.
Волокно диаметром 10 нм может подвергаться дальнейшей конденсации с образованием структур более высокого порядка. При этом нуклеосомы, по всей видимости, образуют соленоид — структуру диаметром 30 нм:
Структура хроматина с разной степенью конденсации. В нижней части рисунка представлен хроматин, находящийся в растянутой форме; он имеет вид нити с нанизанными на нее бусинками. Далее изображен хроматин в частично конденсированной форме, представляющий собой волокно диаметром 10 нм.
В верхней части рисунка представлен хроматин в наиболее конденсированном состоянии, когда волокно диаметром 10 нм образует соленоид диаметром 30 нм.
Обратите внимание на взаимодействие молекул гистона Н1, связанных с каждой нуклеосомой, которое способствует конденсации волокна диаметром 10 нм в более плотную структуру
В результате взаимодействия ДНК с гистонами сегмент двойной спирали ДНК из 168 пар оснований со средним диаметром 2 нм и длиной 57 нм превращается в спираль диаметром 10 нм и длиной 5 нм.
При последующем сжатии этой спирали до волокна диаметром 30 нм степень конденсации увеличивается еще в шесть раз. Таким образом, упаковка дуплекса ДНК с пятью гистонами приводит к 50-кратной конденсации ДНК.
Однако даже столь высокая степень конденсации не может объяснить почти 5000-кратное уплотнение ДНК в метафазной хромосоме.
Эукариотический хроматин содержит и другие белки, которые обычно называют негистоновыми. Некоторые из них, например ферменты, необходимые для репликации и экспрессии ДНК, могут связываться с хроматином временно. Белки, принимающие участие в различных процессах регуляции, связываются с ДНК только в специфических тканях или на определенных стадиях дифференциации.
Сегодня пришли новые технологии и методы, благодаря чему микроскопия в биологии стала трехмерной. Появилась возможность рассмотреть хромосому в интерфазном ядре и получить информацию о локализации в нем сразу всех хромосом человека .
Для этого широко применяют гибридизацию in situ (FISH) ДНК индивидуальных хромосом, меченной флуоресцентными красителями, с ДНК интерфазного ядра. Затем с помощью лазерного сканирующего микроскопа получают серию оптических срезов ядра, на которых зарегистрированы интересующие исследователя сигналы.
Такие оптические срезы можно рассматривать отдельно, использовать для создания ортогональных проекций или для реконструкции трехмерной организации клеточного ядра.
Хромосомы прокариот
Насколько известно, в упаковке прокариотической геномной ДНК участвуют только два или три белка. О природе взаимодействия этих белков с ДНК и о структуре конденсированного комплекса белокнуклеиновая кислота известно немного. У Е.
coli, по-видимому, существует лишь один белок или один класс ДНК-связывающих белков, называемых HU-белками; по своему размеру, содержанию лизина и аргинина, антигенным свойствам они сходны с эукариотическим гистоном Н2А. Другой белок, белок II, обнаруженный у Е.
coli и цианобактерий, по повышенному содержанию лизина и ДНК-связывающим свойствам также напоминает эукариотический гистон. Белки HU и II обнаружены в количествах, достаточных для образования комплекса по крайней мере с половиной ДНК Е.
coli и, по-видимому, совместно с полиаминами и еще неизвестными нам белками могут осуществлять те же самые функции при конденсации и упаковке ДНК, что и пять эукариотических гистонов.
Митоз
Митоз, или непрямое деление, – основной способ размножения эукариотических клеток, обусловливающий, в частности, возможность увеличения их биомассы, рост и регенерацию. Митоз состоит из четырех фаз.
Первая – профаза – характеризуется началом цикла компактизации хромосом, который продолжается в течение всей этой фазы.
Вследствие этого хромосомы становятся видимыми под микроскопом, причем уже в средней профазе митоза они представляются двойными структурами – сестринскими хроматидами, которые являются таковыми, пока удерживаются центромерой вместе. К концу профазы исчезают ядрышко и ядерная мембрана.
Вторая –метафаза. Процесс компактизации хромосом продолжается и ведет к еще большему укорочению их длины. Хромосомы выстраиваются по экватору клетки.
Хроматиды соединены между собой между собой в центромере, называемой также первичной перетяжкой. Появляются нити митотического веретена, которые присоединяются к ценромерам.
Каждая ценромера испытывает напряжение, поскольку нити веретена тянут ее к противоположным полюсам.
Полюса клетки формируются специальными органеллами – центросомами.
Третья – анафаза – начинается с разрыва ценромеры, в результате чего сестринские хроматиды расходятся к разным полюсам клетки. С этого момента каждая пара сестринских хроматид получает название дочерних хромосом.
Четвертая – телофаза. Хромосомы достигают полюсов клетки, появляются ядерная мембрана, ядрышко. Происходят декомпактизация хромосом и восстановление структуры интерфазного ядра. Заканчивается митоз делением цитоплазмы и в типичных случаях – восстановлением исходной биомассы дочерних клеток.
Биологическая роль митоза состоит в обеспечении идентичной генетической информацией двух дочерних клеток.
Это достижимо только благодаря циклу компактизации – декомпактизации, который и позволяет распределить наследственные молекулы в минимальном объеме митотических хромосом.
В противном случае, учитывая размеры клетки (десятки или сотни кубических микрометров) и длину декомпактизованной хромосомы (сантиметры), каждое клеточное деление сопровождалось бы хаотичным переплетением хромосомного материала.
В эволюции эукариотических клеток, видимо, это обстоятельство и послужило причиной становления столь сложного генетического процесса, как митоз.
Каждая хромосома индивидуальна, т.е. характеризуется свойственными только ей размерами, формой и положением центромеры. В клетках тела организмов, размножающихся половым путём, любая хромосома представлена двумя копиями, или гомологами .
При образовании половых клеток в мейозев каждую из них попадает одна из двух гомологичных хромосом . При оплодотворении парность гомологичных хромосом восстанавливается: одна хромосома каждой пары отцовская, другая – материнская.
Совокупность признаков хромосомного набора (число хромосом, их размер и форма) постоянна для клеток каждого вида и называется его кариотипом. В кариотипе различают пару определяющих пол организма половых хромосом и все остальные хромосомы – аутосомы.
Изучением поведения хромосом в митозе и мейозе, а также роли хромосом, особенно половых, при передаче признаков от одного поколения к другому привело к созданию в нач. 20 в.
хромосомной теории наследственности и до настоящего времени исследуется огромным количеством как цитогенетиков, так и других ученых, включая и физиков. Как уже было сказано, хромосомой часто называют генетический материал бактерий и вирусов, хотя его строение отличается от хромосом эукариотических организмов.
Источник: https://stydopedia.ru/1x71c.html
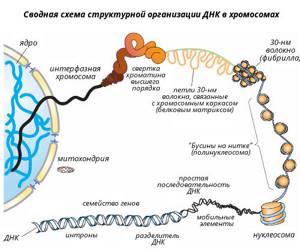
Структура хроматина. Упаковка ДНК
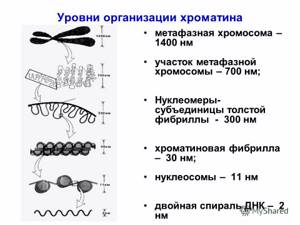
Первый уровень упаковки нитей ДНК- нуклеосомный– связан с участием в этом процессе специальных ядерных белков – гистонов.Во всех эукариотических клетках выделено пять фракций гистонов (Н1, Н2А, Н2B, H3, H4).
Группа из 8 молекул гистонов (по две молекулы каждого из гистонов Н2А, H2B, H3,H4), образуют глобулу, вокруг которой по спирали закручивается участок ДНК размером в 146 пар нуклеотидов. Эти глобулы «бусинки» были открыты в 1974 году и названы нуклеосомами.
Нуклеосомный уровень укладки хроматина позволяет «сжать» молекулу ДНК примерно в 6-7 раз.
Второй уровень упаковки нитей ДНК –нуклеомерный.
несколько нуклеосом сближаются, формируют компактные группы – нуклеомеры,которые, в свою очередь образуют суперспираль.
Такая компактная нить ДНК имеет диаметр 30 нм и получила наименование – хроматиновая фибрилла. При этом происходит еще более чем 40 – кратное сжатие ДНК.
Третий уровень – петлевой или доменный. На этом уровне хроматиновые фибриллы образуют систему петель (доменов) диаметром 300 нм, в каждой из которых находится несколько генов. Это приводит к еще 600-кратному сжатию ДНК
Четвертый уровень–хромомерный Происходит переход от спирального типа укладки ДНК, состоящих из петель хроматиновых 30-нм фибрилл, к образованию компактных глобулярных структур типа хромомеров, имеющих уже размеры 0,1-0,2 мкм..Степень сжатия – несколько тысяч раз.
Пятый уровень – стадия конденсированной метафазной хромосомы
Это уровень сверхкомпактного состояния хроматина. Видны только в период митоза. Неактивны – считывание информации не происходит. К кинетохорам метафазных хромосом крепятся микротрубочки веретена деления.
Нуклеосомы (первый уровень)
Хиазмы Нуклеомер(второй уровень)
Петлевой и хромомерный уровень упаковки ДНК.(третий и четвертый уровни) Видны петли ДНК и плотные (хромомерные) участки между ними
(гигантская интерфазная хромосома в яйцеклетке птиц- типа «ламповой щетки») Стрелки указывают на участки перекреста (кроссинговера) гомологичных хромосом (хиазмы)
Структура гена. Транскрипция
Ген – структурная единица генетической информации, закодированная в определенной последовательности нуклеотидов участка ДНК.
Генотип – совокупность гаплоидного набора всех генов данной клетки или организма.
В дальнейших исследованиях было установлено, что:
– ген дискретен и делим;
– ген имеет очень сложную структуру;
– информация записана в гене посредством универсального генетического кода;
– в геноме имеются разные типы (группы) генов;
– наименьшей мутирующей единицей гена является пара нуклеотидов – т.н. однонуклеотидные замены (SNP – single nucleotide polymorphisms).
Гены состоят из двух основных частей: регуляторной и кодирующей.
Кодирующая часть гена имеет мозаичную структуру и состоит из функционально разных участков – экзонов и интронов.
С одних – экзонов в дальнейшем считывается информация о белке, в других – интронахне закодирована информация о белке. Объем, занимаемый интронными участками гена, значительно превышает экзонные.
Каждый структурный ген имеет свой промотор
Промотор – это небольшой участок ДНК, расположенный перед структурной частью гена и предназначенный для связывания РНК-полимеразы и белков-регуляторов перед началом синтеза РНК. У высших эукариот в состав промотора входят многократно повторяющиеся последовательности нуклеотидов: ТАТА, ЦААТ, ГЦ
Регуляторные участки гена эукариот, с которыми связываются факторы транскрипции, располагаются за сотни пар нуклеотидов от участка промотора активируемого гена- они получили название – энхансеры.
Каждый ген или группа генов отделены друг от друга специальным участками ДНК, называемые спейсерами.
Схема организации типичного гена эукариот и его регуляторные элементы:
1 – экзоны; 2 – интроны; 3 – промотор; 4 – РНК-полимераза II; 5 – главные факторы транскрипции; 6 – регуляторные белки; 7 – энхансеры (регуляторные участки гена); 8 – спейсерные участки; 9 – ТАТА блок – инициирующая часть промотора; стрелка – начало процесса транскрипции
Источник: https://infopedia.su/10x89a7.html
Маркируем гены, пакуем ДНК: ученые исследовали способы упаковки ДНК
Ученые МГУ имени М.В. Ломоносова создали метод маркировки работающих генов, основываясь на различиях активных и неактивных участков хромосом во время репликации из ДНК. Статья опубликована в журнале Current Biology.
Совместно со своими коллегами из США российские ученые исследовали способы упаковки ДНК в клеточном ядре и их изменение в процессе ее репликативного синтеза. Считается, что характер упаковки ДНК оказывает существенное влияние на работу генов и является одним из механизмов эпигенетического контроля генной экспрессии.
«Эпигенетический контроль генной экспрессии выражается в том, что, хотя все клетки организма имеют одинаковую генетическую информацию, не все гены работают в данном отдельно взятом типе клеток. Именно набор работающих генов определяет судьбу клетки.
Существуют несколько клеточных механизмов, которые помогают клетке запомнить, какие гены должны в ней работать, а какие — нет.
Это и есть эпигенетический контроль, представляющий собой память клетки о том, кто она и благодаря работе каких генов она такой является, — рассказывает один из авторов статьи, заведующий отделом электронной микроскопии НИИ физико-химической биологии имени А.Н.
Белозерского, доктор биологических наук Игорь Киреев. — Нас интересовали так называемые высшие уровни структурной организации хромосом, формирующиеся в результате серии последовательных этапов упаковки нити ДНК».
ДНК в клетке существует в виде комплекса с белками — хроматина. Начальные этапы компактизации хроматина достаточно хорошо исследованы — это нуклеосомы, белковые глобулы размером около 10 нм, состоящие из восьми молекул белков-гистонов, на которые наматывается ДНК.
Затем цепочка нуклеосом укладывается непонятным пока образом в более толстые фибриллы, хромонемы, в результате чего достигается очень высокая степень компактизации.
Так, длина максимально компактизованной митотической хромосомы в 20 000 раз меньше длины уложенной в ней ДНК.
Репликация — это процесс синтеза дочерней молекулы ДНК на матрице родительской молекулы, а транскрипция — процесс синтеза РНК с использованием ДНК в качестве матрицы.
ДНК в составе хромосом чрезвычайно плотно и сложно упакована, и традиционно считалось, что для осуществления процессов матричного синтеза (транскрипция и репликация) эта упаковка мешает и должна быть нарушена в масштабе довольно больших по размеру хроматиновых доменов.
Идентифицировать эти домены и проанализировать их структурную организацию с высоким пространственным разрешением, не нарушив при этом их естественной структуры, было достаточно сложно.
«Мы предложили метод маркировки работающих генов, основываясь на различиях активных и неактивных хромосомных участков во время репликации из ДНК.
Так, активно работающий участок, так называемый эухроматин, реплицируется в самом начале синтетического периода клеточного цикла, а «молчащий» — гетерохроматин — во второй его половине, — комментирует Игорь Киреев.
— Наш метод позволил осуществлять недеструктивные (неразрушающие) исследования хроматина.
Он основан на комбинации мечения новосинтезированной (дочерней) ДНК методами click-химии с последующей детекций продуктов реакции методами корреляционной флуоресцентной микроскопии с суперразрешением и иммуноэлектронной микроскопии. Иными словами, одну и ту же молекулу в данной клетке мы можем видеть и в оптический, и в электронный микроскоп».
Используя данный подход, ученые сделали два неожиданных наблюдения. Во-первых, «работающий» хроматин, вопреки традиционным представлениям, сохраняет весьма высокую степень упаковки, поскольку он представлен высоко структурированными хроматиновыми фибриллами высшего порядка — хромонемами.
Во-вторых, оказалось, что ДНК в составе хромонем обладает высокой структурной пластичностью, то есть способна как бы «перетекать» из одного участка хромонемы в соседний. При этом общая плотная структура хромонемы не изменяется.
Эти наблюдения не укладываются в рамки существующих теорий о пространственной организации хромосом, но в то же время позволяют высказать новые гипотезы о механизмах передачи эпигенетической информации в процессе клеточного деления.
«Мы сделали предположение о том, что в новой дочерней клетке структуры хроматина могут перемещаться внутри нее, а не являться фиксированными, взаимодействуя с той ДНК, которая еще не удваивалась и «помнит все», а также содержит необходимые молекулярные компоненты для восстановления утраченной эпигенетической информации», — продолжает Игорь Киреев.
Другой озвученный в статье вывод состоит в том, что структурная организация генома не является жесткой иерархией. Конечно, есть некие последовательные уровни организации ДНК.
Раньше думали, что должна быть четко зафиксированная система, которая однозначно приводит к формированию хромосом.
Сейчас оказывается, что все может быть по-другому: есть некие принципы построения, но внутри заданных границ ДНК обладает некоторой свободой и пластичностью.
«Дальнейшее развитие исследований состоит, во-первых, в переходе к анализу индивидуальных хромосомных локусов, которые мы планируем маркировать (выделять) на оптическом и электронно-микроскопическом уровнях при помощи технологии TALE, а во-вторых, в разработке еще более нативных методов in vivo мечения, совместимых с такими передовыми технологиями, как криоэлектронная микроскопия», — делится планами на будущее Игорь Киреев.
Ученые надеются вплотную приблизиться к расшифровке принципов пространственной организации ДНК, используя прямые методы анализа при помощи визуализации способов упаковки хроматина с высоким разрешением.
В практическом плане исследования позволят выяснить структурные аспекты эпигенетического контроля генной экспрессии и, возможно, подсказать пути его регулирования, что позволит разрабатывать более эффективные подходы для терапевтических воздействий, например при борьбе с раком и старением — состояниями, в которых очень ярко выражена «эпигенетическая» компонента.
Источник: https://indicator.ru/article/2016/11/07/uchenye-mgu-issledovali-sposoby-upakovki-dnk-v-kletochnom-yadre/