В этой статье мы разберемся, что такое мейоз и через какие фазы он проходит. Поймем какой хромосомный набор на каждом этапе такого деления и что обозначают все эти n и c. А самое главное — какое биологическое значение у мейоза. В конце сравним его с митозом, выявим сходства и различия между ними.
Что же такое мейоз?
Мейоз — это способ деления клетки. Его еще называют редукционным делением, потому что из одной диплоидной клетки получается четыре гаплоидных, то есть происходит уменьшение хромосом в два раза.
Какие клетки могут так делиться? Эукариотические, но не все, а только избранные. Прежде всего, это предшественники половых клеток человека — сперматоциты и овоциты (или ооциты). Ещё таким способом образуются споры у высших растений.
Мейоз проходит через два этапа — редукционный и эквационный. Каждый из них разделяется на четыре фазы: профаза, метафаза, анафаза и телофаза. Но перед тем , как рассмотреть этапы и фазы давайте разберемся из чего складывается хромосомный набор.
Хромосомный набор
Начнем с хромосомы. Представьте себе мешок с картошкой. Вот хромосома — это такой мешок, только вместо картошки в ней длинная молекула ДНК, которая связана с белками — гистонами и негистонами.
Всего у нас 46 хромосом или 23 пары. Почему пары? Дело в том, что у каждой хромосомы есть своя сестричка — двойняшка (гомолог). Вроде они и очень похожи, но разница есть. Они содержат похожие молекулы ДНК, но не такие же! Гомологичные хромосомы могут содержать немного разные нуклеотидные последовательности, а значит по-разному проявляют признаки.
Когда у каждой хромосомы есть своя пара, то это диплоидный набор — 46 хромосом. Если пары нет, то это гаплоидный набор — 23 хромосомы.
Рисунок. Гомологичные хромосомы
n — это число хромосом. У каждой есть своя пара, значит всего 2n.
c — это число молекул ДНК, в одной хромосоме одна молекула. Всего молекул = 2c
Редукционный этап или первое деление мейоза
Его суть — изменение числа хромосом внутри клетки. То есть из одной диплоидной (2n4c) клетки получаем две гаплоидных (1n2c). Так стоп, откуда 4c? До этого же было 2n2c. Ах да… Сейчас разберемся.
Интерфаза
Перед вступлением в мейоз клетка проходит через интерфазу. Ей нужно подготовиться к делению — запасти энергетических субстратов (АТФ), синтезировать необходимые белки и удвоить количество молекул ДНК. Еще в интерфазу происходит удваивание центромер.
Рисунок. Хромосомный набор в интерфазу
В интерфазу произошла репликация ДНК — образовалась идентичная цепь. Но эти две цепи, или хроматиды, связаны между собой при помощи центромеры, значит количество хромосом такое же. Итого набор — 2n4c
Ну вы ведь понимаете, что таким образом реплицируются все 46 хромосом. Просто удобнее показать на паре. Помните, что все 23 пар вступают в мейоз, а не только одна. После репликации начинается собственно мейоз, а именно его первая фаза:
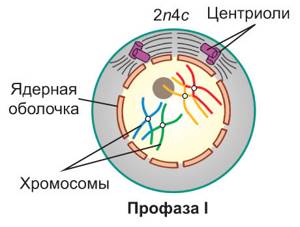
Профаза мейоза I
В отличие от митоза состоит из пяти стадий: лептотена, зиготена, пахитена диплотена и диакинез. Она более длительная и здесь протекают важные процессы: конъюгация и кроссинговер. Еще в эту фазу растворяется ядерная оболочка и формируется веретено деления, подробнее об этом ниже.
Лептотена
Какая основная задача у клетки? Правильно, передать генетический материал своим потомкам.
Поэтому она начинает упаковывать молекулы ДНК как можно плотнее, она собирает чемодан, ведь не хочет ничего не потерять в пути. Этот процесс называется спирализация или конденсация хромосомы.
Клетка так старается, что невидимые раньше в микроскоп хромосомы становятся видимыми. Они похожи на длинные и тонкие нити.
Зиготена
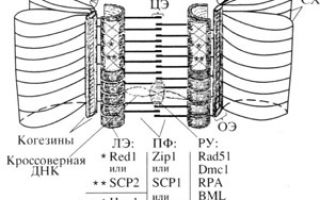
Здесь происходит конъюгация хромосом — их сближение с образованием бивалентов. Связь обеспечивает синаптонемальный комплекс — он удерживает гомологичные хромосомы рядом это необходимо для запуска кроссинговера на следующем этапе.
Схема. Образование бивалентов.
Связи между хромосомами могут иметь разный вид, но они должны быть. Если в клетке останутся хромосомы, которые не сблизились, то она запускает апотоз и погибает. Клетка — с заботой о будущих поколениях!
Пахитена
Начинается с еще большей конденсации хромосом, они становятся короче и толще. Но в местах образования синаптонемальных комплексов происходит частичное раскручивание (деконденсация) хромосом.
Все это для начала кроссинговера — обмена участками ДНК у гомологичных хромосом. Обмен обеспечивает перекомбинацию генетического материала. Если бы мы могли рассоединить хромосомы сразу после кроссинговера, то увидели примерно такую картину:
Схема. Кроссинговер.
Это лишь схематичное изображение, перекресты могут происходить в самых разных местах , что дает огромную генетическую вариабельность.
В конце пахитены мостики между хромосомами разрушаются, они начинают отдаляться друг от друга.
Диплотена
Хромосомы расходятся в области центромер, но остаются связаны между собой в местах кроссинговера — перекрестах или хиазмах. В микроскоп можно увидеть все четыре хроматиды, так сильно они упаковались (спирализовались).
Диакинез
Гомологичные хромосомы расходятся, формируется веретено деления и исчезает ядерная оболочка. Этим завершается профаза мейоза I. Вид клетки примерно такой:
Схема. Конец профазы мейоза I
2n4c
Хромосомный набор в конце профазы I
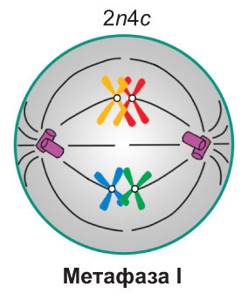
Метафаза мейоза I
В этой фазе заканчивается образование веретена деления. Нити веретена прикрепляются к центромерам и начинают притягивать хромосомы, из-за этого они располагаются на экваторе клетки.
Схема. Клетка в метафазу I
2n4c
Набор в метафазе I
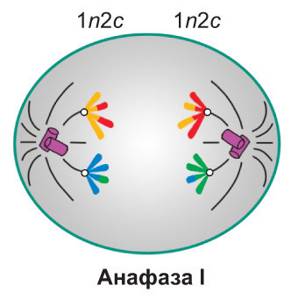
Анафаза мейоза I
Нити веретена деления продолжают тянуть хромосомы на себя — они расходятся к полюсам клетки. На полюсах клетки располагается по 23 хромосомы, но они все еще состоят из двух нитей ДНК.
Схема. Анафаза мейоза I
n2c у полюсов, но вся клетка 2n4c
Набор в анафазу I
Телофаза мейоза I
Завершение редукционного деления. Появляется ядерная оболочка, которая окружает хромосомы. Затем возле ядер появляется перетяжка, которая делит клетку на две части. Образуются две гаплоидные клетки.
Схема. Конец первого деления мейоза
Образование двух гаплоидных клеток — n2c
Результат редукционного деления
Второй этап мейоза — эквационный
Начинается сразу же после первого. Эквация — это уравнивание. Так что задача клетки на этом этапе — сделать так, чтобы в одной хромосоме была одна молекула ДНК.
Он похож на митоз, здесь к полюсам клетки отправятся хроматиды, а не целые хромосомы и мы получим из каждой клетки по две — с набором nc.
Протекает он через такие же фазы, но с одним исключением. Здесь не будет интерфазы — клетка уже готова к делению, она запасла энергетические субстраты и белки ещё перед началом первого деления. Поэтому сразу начинается профаза II.
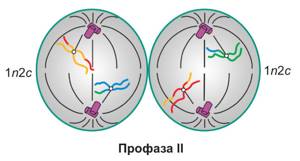
Профаза мейоза II
Клетка уже сделала свою работу — упаковала генетический материал как можно лучше. Ей ничего не нужно делать, ну почти. Разве что растворить ядерные оболочки и достроить веретено деления. Этим она и займется.
Схема. Профаза II
Вы конечно понимаете, что вторая клетка идет по такому же пути. Просто мне лень рисовать сразу две.
n2c
Набор в профазу II
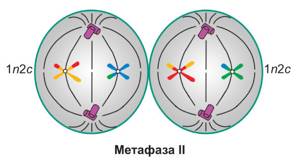
Метафаза мейоза II
Прикрепление нитей веретена деления к центриолям — хромосомы снова на экваторе клетки.
Схема. Клетка в метафазу II
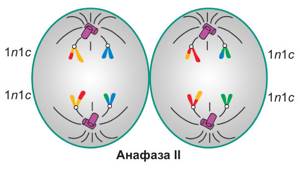
Анафаза мейоза II
Торжественный момент — сейчас наши хроматиды станут полноценными хромосомами. Каждая разойдется к своему полюсу.
Схема. Клетка в анафазу мейоза II
Поздравляем, ох уж эти хроматиды, они так быстро растут…
У полюсов — nc, всего 2n2c, так как каждая хроматида теперь — это полноценная хромосома.
Набор в анафазу II
Телофаза мейоза II
Вокруг хромосом формируются ядерные мембраны, появляется перетяжка и делит клетку на две.
Схема. Вот и закончился мейоз
Вторая клетка прошла через такое же деление. Всего из одной диплоидной клетки 2n2c получилось четыре гаплоидных с набором nc.
Четыре клетки с хромосомным набором — nc
Результат мейоза
Биологическое значение мейоза
1)Передать свой генетический материал будущим поколениям.
2)Поддержать диплоидный набор хромосом у организма. В конце мейоза формируются гаплоидные клетки, которые после оплодотворения образуют диплоидный набор.
3)Мейоз обеспечивает не только передачу генетической информации, но и ее преобразование — основа изменчивости. Кроссинговер обеспечивает взаимный обмен у гомологичных хромосом. В анафазу I к полюсам клетки независимо расходятся гомологичные хромосомы, а в анафазу II — хроматиды. Так формируются уникальные комбинации генов.
Сходства и различия мейоза и митоза
Таблица. Сходства и различия мейоза и митоза
Биология. 11 класс
Мейоз — особый способ деления эукариотических клеток, в результате которого из одной материнской клетки образуются четыре дочерние с уменьшенным в 2 раза набором хромосом. Если в мейоз вступает диплоидная клетка (2n4c), то образуются четыре гаплоидные клетки (1n1c). Клетки с гаплоидным набором хромосом не способны делиться мейозом.
Мейоз представляет собой два последовательных деления — мейоз I и мейоз II. Важно отметить, что репликация ДНК предшествует только первому мейотическому делению.
Между мейозом I и мейозом II удвоения ДНК не происходит. Каждое из двух делений обычно включает профазу, метафазу, анафазу и телофазу. Рассмотрим процесс мейотического деления диплоидной клетки.
Первое деление мейоза осуществляется следующим образом (табл. 18.1)
Таблица 18.1. Первое деление мейоза (мейоз I)
| Схема | Фаза и процессы, происходящие в ней |
| Профаза I. В ядре начинается спирализация хроматина. *Длинные тонкие хроматиновые нити постепенно укорачиваются и при этом утолщаются, и формирующиеся хромосомы становятся четко различимыми в световой микроскоп.* Каждая хромосома состоит из двух хроматид. Содержание наследственного материала в клетке выражается записью 2n4c. Гомологичные хромосомы попарно сближаются и соединяются друг с другом *сначала в нескольких участках, а затем по всей своей длине*. Этот процесс называется конъюгацией гомологичных хромосом, *а образовавшиеся хромосомные пары — бивалентами. В ходе конъюгации хромосомы тесно соприкасаются. В некоторых точках соприкосновения, называемых хиазмами, вследствие разрыва и последующего воссоединения молекул ДНК,* между хроматидами гомологичных хромосом может происходить обмен соответствующими участками — кроссингóвер. *Далее гомологичные хромосомы в составе каждого бивалента начинают отталкиваться друг от друга и в результате остаются связанными только в областях хиазм.* Одновременно с этим в клетке начинается образование веретена деления. К концу профазы I исчезают ядрышки и распадается ядерная оболочка | |
| Метафаза I. Завершается формирование веретена деления. Спирализация хромосом достигает максимума. *Биваленты* располагаются в центральной части клетки, *образуя метафазную пластинку. Каждый бивалент ориентирован таким образом, что центромеры гомологичных хромосом находятся по разные стороны от экваториальной плоскости клетки, на одинаковом расстоянии от нее.* При этом нити веретена деления, идущие от противоположных полюсов клетки, прикреплены к центромерам разных гомологичных хромосом. Таким образом, в составе каждого *бивалента* одна из хромосом оказывается связанной с одним полюсом клетки, а другая — с противоположным | |
| Анафаза I. *Каждый бивалент распадается на две хромосомы, причем их* центромеры, в отличие от митоза, не разделяются надвое. Поэтому нити веретена деления растягивают к противоположным полюсам клетки не сестринские хроматиды, как при митозе, а гомологичные хромосомы. Следовательно, диплоидный набор 2n4c разделяется на два гаплоидных набора, которые впоследствии попадут в разные дочерние клетки. Каждая хромосома состоит из двух хроматид, которые в результате кроссинговера уже не идентичны друг другу. Таким образом, в конце анафазы I набор хромосом и хроматид у каждого полюса делящейся клетки составляет 1n2c | |
| Телофаза I. Веретено деления разрушается. Происходит деспирализация хромосом и формирование двух ядер. Далее клетка разделяется на две дочерние. Они имеют гаплоидный набор хромосом, каждая хромосома состоит из двух хроматид (1n2c) |
Промежуток времени между первым и вторым делениями мейоза обычно очень короткий. В этот период, как уже отмечалось, не осуществляется репликация ДНК. Каждая из двух клеток, образовавшихся в результате мейоза I, вступает в мейоз II. Это деление протекает аналогично митозу (табл. 18.2).
*У некоторых организмов перерыв между мейозом I и мейозом II — так называемый интеркинез вообще отсутствует. При этом клетки, завершившие телофазу I, сразу же переходят в профазу II. Более того, могут выпадать и эти фазы.
Так, у большинства растений, клетки, делящиеся мейозом, после окончания анафазы I вступают непосредственно в метафазу II.
В этом случае переход от первого мейотического деления ко второму не сопровождается деспирализацией хромосом, формированием двух ядер и разделением материнской клетки на две дочерние.*
Таблица 18.2. Второе деление мейоза (мейоз II)
| Схема | Фаза и процессы, происходящие в ней |
| Профаза II. В результате спирализации хроматина формируются двухроматидные хромосомы (набор 1n2c). В это же время начинается образование веретена деления. После распада ядерной оболочки отдельные хромосомы беспорядочно располагаются в гиалоплазме | |
| Метафаза II. Завершается формирование веретена деления. Хромосомы, достигшие максимальной спирализации, выстраиваются в центральной части клетки, *формируя метафазную пластинку*. Нити веретена деления связывают центромеру каждой хромосомы с двумя противоположными полюсами | |
| Анафаза II. Происходит разделение центромер. Сестринские хроматиды (теперь уже дочерние хромосомы) растягиваются к разным полюсам клетки. В конце анафазы II набор хромосом и хроматид у каждого полюса составляет 1n1c | |
| Телофаза II. Хромосомы деспирализуются, формируются ядра, и происходит разделение клеток. При этом образуются четыре дочерние клетки, имеющие набор 1n1c |
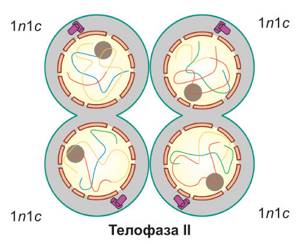
Таким образом, в результате первого деления мейоза исходная материнская клетка (2n4c) разделилась на две дочерние, имеющие гаплоидный набор хромосом.
*Следовательно, в мейозе I произошла редукция (уменьшение) числа хромосом в 2 раза. Поэтому первое деление называют редукционным.
В образовавшихся гаплоидных клетках каждая хромосома представлена двумя хроматидами (1n2c).
В результате второго деления, протекающего по типу митоза, набор хромосом (1n) не изменяется. Это деление мейоза называется эквационным (от лат. aequatio — уравнивание).
Эквационное деление, в отличие от редукционного, сопровождается расхождением сестринских хроматид.
Поэтому итогом второго деления является образование четырех гаплоидных клеток с однохроматидными хромосомами (1n1c).*
Мейоз — более длительный процесс, чем митоз. Например, у ржи он протекает более 2 сут, у репчатого лука — около 4 сут. Мейотическое деление клеток мыши занимает примерно неделю. Мейоз, протекающий при образовании сперматозоидов человека, длится приблизительно 25 дней. Самой продолжительной стадией мейоза является профаза I.
*Как и в случае митоза, правильное протекание мейоза может быть нарушено действием определенных внешних или внутриклеточных факторов. Последствия таких воздействий могут быть разными, вплоть до образования нежизнеспособных дочерних клеток.
Наиболее часто встречающейся патологией мейоза является нерасхождение хромосом в анафазе I. Оно происходит вследствие нарушения разделения бивалентов и выражается в том, что обе гомологичные хромосомы перемещаются к одному из полюсов клетки.
Нерасхождение может наблюдаться и на стадии анафазы II. При этом к одному и тому же полюсу отходят две сестринские хроматиды (дочерние хромосомы). Как в первом, так и во втором случае результатом нерасхождения хромосом является образование генетически неравноценных клеток.
В одних клетках наблюдается избыток хромосом (1n + 1), а в других — недостаток (1n — 1).*
Биологическое значение мейоза. У животных и человека путем мейоза образуются гаметы — гаплоидные половые клетки. В результате последующего оплодотворения формируется зигота с двойным набором хромосом, из которой развивается новый организм.
Он является диплоидным, как и его родители, а значит, сохраняет свойственный данному виду организмов кариотип. Без мейоза, приводящего к уменьшению набора хромосом в 2 раза, половое размножение сопровождалось бы удвоением числа хромосом в каждом новом поколении.
У растений, многих водорослей и грибов мейоз приводит к формированию спор, с помощью которых осуществляется бесполое размножение.
Кроссинговер, происходящий в профазе I, приводит к перекомбинации наследственного материала между гомологичными хромосомами. В анафазе I гомологичные хромосомы каждой пары случайным образом, независимо от других пар, расходятся к разным полюсам клетки.
В анафазе II то же самое происходит с сестринскими хроматидами.
Все эти процессы являются важными источниками комбинативной изменчивости (будет рассмотрена подробно в главе 6), обеспечивающей появление разнообразного потомства как при половом размножении, так и при размножении спорами.
Мейоз кратко и понятно
- Мейоз – осуществляется в клетках организмов, размножающихся половым путем.
- Биологический смысл явления определяется новым набором признаков у потомков.
- В данной работе рассмотрим сущность этого процесса и для наглядности представим его на рисунке, посмотрим последовательность и продолжительность деления половых клеток, а так же узнаем, в чем сходство и отличие митоза и мейоза.
Что такое мейоз
Процесс, сопровождающийся образованием четырех клеток с одинарным хромосомным набором из одной исходной.
Генетическая информация каждой новообразованной соответствует половине набора соматической клетки.
Фазы мейоза
Мейотичекое деление включает два этапа, состоящие из четырех фаз каждое.
Первое деление
Включает профазу I, метафазу I, анафазу I и телофазу I.
Профаза I
На данном этапе образуются две клетки с половинным набором генетической информации. Профаза первого деления включает несколько стадий. Ей предшествует предмейотическая интерфаза, во время которой идет репликация ДНК.
Затем происходит конденсация, образование длинных тонких нитей с протеиновой осью во время лептотены. Данная нить прикрепляется к мембране ядра с помощью концевых расширений – прикрепительных дисков. Половинки удвоенных хромосом (хроматиды) еще не различимы. При исследовании имеют вид монолитных структур.
Далее наступает стадия зиготены. Гомологи сливаются с образованием бивалентов, число которых соответствует одинарному числу хромосом.
Процесс конъюгации (соединения) осуществляется между парными, сходными в генетическом и морфологическом аспекте. Причем взаимодействие начинается с концов, распространяясь вдоль тел хромосом.
Комплекс из гомологов, связанных белковым компонентом – бивалент или тетрада.
Спирализация происходит во время стадии толстых нитей – пахитены. Здесь уже удвоение ДНК выполнено полностью, начинается кроссинговер. Это обмен участками гомологов. В результате формируются сцепленные гены с новой генетической информацией. Параллельно протекает транскрипция. Плотные участки ДНК – хромомеры активируются, что приводит к изменению структуры хромосом по типу «ламповых щеток».
Гомологичные хромосомы конденсируются, укорачиваются, расходятся (исключая точки соединения хиазмы). Это стадия в биологии диплотена или диктиотена. Хромосомы на данном этапе богаты РНК, которая синтезируется на этих же участках. По свойствам последняя близка к информационной.
Наконец, биваленты расходятся к периферии ядра. Последние укорачиваются, теряют ядрышки, становятся компактными, не связанными с ядерной оболочкой. Это процесс носит название диакинеза (перехода к делению клетки).
Виды и формы бесполого размножения
Далее биваленты перемещаются к центральной оси клетки. От каждой центромеры отходят веретена деления, каждая центромера равноудалена от обоих полюсов. Небольшие по амплитуде движения нитей удерживают их в данном положении.
Анафаза I
Хромосомы, построенные из двух хроматид, расходятся. Происходит перекомбинация с уменьшением генетического разнообразия (в связи с отсутствием в наборе генов, расположенных в локусах (участках) гомологов).
Телофаза I
Суть фазы состоит в расхождении хроматид с их центромерами к противоположным участкам клетки. В животной клетке происходит цитоплазматическое деление, в растительной – образование клеточной стенки.
Второе деление
После интерфазы первого деления клетка готова ко второму этапу.
Профаза II
Чем длиннее телофаза, тем короче длительность профазы. Хроматиды выстраиваются вдоль клетки, образуя своими осями прямой угол относительно нитей первого мейотического деления. В эту стадию они укорачиваются и утолщаются, ядрышки подвергаются распаду.
Центромеры вновь расположены в экваториальной плоскости.
Анафаза II
Хроматиды отделяются друг от друга, перемещаясь к полюсам. Теперь они носят название хромосом.
Телофаза II
Деспирализация, растяжение образованных хромосом, исчезновение веретена деления, удвоение центриолей. Гаплоидное ядро окружается ядерной мембраной. Формируются четыре новые клетки.
Таблица сравнения митоза и мейоза
Кратко и понятно особенности и отличия представлены в таблице.
| Характеристики | Мейотическое деление | Митотическое деление |
| Число делений | осуществляется в два этапа | осуществляется в один этап |
| Метафаза | после удвоения хромосомы расположены по центральной оси клетки парами | после удвоения хромосомы расположены по центральной оси клетки одиночно |
| Слияние | есть | нет |
| Кроссинговер | есть | нет |
| Интерфаза | нет удвоения ДНК в интерфазу II | перед делением характерно удвоение ДНК |
| Итог деления | гаметы | соматические |
| Локализация | в зреющих гаметах | в соматических клетках |
| Путь воспроизведения | половой | бесполый |
Представленные данные – схема отличий, а сходства сводятся к одинаковым фазам, редупликации ДНК и спирализации перед началом клеточного цикла.

Какова же роль мейоза:
- Дает новые сочетания генов вследствие кроссинговера.
- Поддерживает комбинативную изменчивость. Мейоз – источник новых признаков в популяции.
- Удерживает постоянное количество хромосом.
Заключение
Мейоз сложный биологический процесс, в ходе которого образуются четыре клетки, с новыми признаками, полученными в результате кроссинговера.
На пути к разгадке тайны мейоза
Появление мейоза – особого варианта клеточного деления, в результате которого число хромосом сокращается вдвое – было одним из важнейших эволюционных «достижений» первых эукариот. Механизм мейоза сложился из комбинации готовых «блоков»: механизмов митоза, рекомбинации и репарации ДНК. Ключевым событием стало формирование синаптонемного комплекса – особой белковой структуры, обеспечивающей попарное соединение и точное «выравнивание» хромосом.
Появление эукариотической клетки было важнейшим эволюционным преобразованием (ароморфозом) в истории земной жизни (см. обзор «Происхождение эукариот» ).
Одним из главных «достижений» древних эукариотических организмов стало возникновение настоящего полового процесса, то есть слияния двух гаплоидных (содержащих одинарный набор хромосом) клеток – гамет в диплоидную (содержащую двойной набор хромосом) клетку – зиготу.
Чтобы жизненный цикл эукариот, обладающих половым процессом, мог продолжаться, должен был развиться механизм, посредством которого из диплоидных клеток снова могли образовываться гаплоидные. Таким механизмом стал мейоз – особый вид клеточного деления, при котором число хромосом в дочерних клетках уменьшается вдвое по сравнению с родительской клеткой.
В статье Ю.Ф.Богданова, крупнейшего специалиста по эволюции мейоза, рассматриваются современные представления о происхождении механизмов мейотического деления у эукариот.
Общепризнано, что мейоз произошел из митоза – «обычного» клеточного деления эукариот, в результате которого число хромосом остается прежним. Происхождение митоза само по себе было замечательным ароморфозом. Мейоз, по сути дела, является модифицированной версией митоза.
Мейоз представляет собой два последовательных деления исходной диплоидной клетки и отличается от митоза только первым своим делением (см. схему).
Отличие состоит в том, что хромосомы вступают в метафазу, соединенные попарно гомолог с гомологом (гомологичными называют хромосомы, содержащие одни и те же гены/локусы и полученные одна от отца, другая от матери).
В метафазе I мейоза центромеры каждой из хромосом «униполярны», т.е. соединены белковыми нитями только с одним из двух полюсов веретена деления .
В ходе митоза (а также в ходе второго деления мейоза) хромосомы вступают в метафазу поодиночке, и нити веретена деления присоединяются к каждой хромосоме с двух сторон.
Именно благодаря указанным особенностям первого деления мейоза и обеспечивается уменьшение числа хромосом: к полюсам клетки расходятся не сестринские хроматиды, а гомологичные хромосомы, по одной из каждой пары (см. схему).
Эти ключевые особенности первого деления мейоза развились на основе одного и того же ароморфоза – возникновения так называемого синаптонемного комплекса (СК).
Механизм мейоза был «собран» из готовых блоков (как это часто бывает в эволюции) – разумеется, с некоторыми модификациями и новшествами.
Одним из этих исходных блоков стал уже имевшийся к тому времени у эукариот механизм митоза, другим – механизм репарации (починки повреждений) ДНК, основанный на гомологичной рекомбинации (см.
: Глазер В.М. Гомологичная генетическая рекомбинация ).
Суть процесса в том, что поврежденный участок одной молекулы ДНК заменяется его неповрежденной копией, взятой из другой (гомологичной) молекулы ДНК.
Чтобы «развести» гомологичные хромосомы к разным полюсам клетки, в ходе мейоза используется веретено деления, «унаследованное» от митоза.
Но чтобы распределение хромосом по дочерним клеткам прошло без ошибок, гомологичные хромосомы необходимо сначала сгруппировать попарно.
Для этого используется механизм гомологичной рекомбинации, точнее, одна из деталей этого механизма – образование так называемого соединения Холлидея.
Починка разрывов ДНК – одна из основ мейоза
Механизмы репарации и рекомбинации ДНК – очень древние, они должны были возникнуть еще на заре жизни.
Жизненный цикл древних эукариот (как и многих прокариот), вероятно, состоял из двух фаз и двух соответствующих типов метаболизма: 1) вегетативная фаза (условия благоприятны, клетки размножаются митозом); 2) фаза споруляции (условия стали неблагоприятными, клетки превращаются в споры). У низших эукариот – например, у дрожжей, – перед споруляцией происходит мейоз, и из получившихся гаплоидных клеток образуются споры.
Становление мейоза в ходе эволюции, так же как и сам мейоз в жизненном цикле современных организмов, начинается с репарации повреждений (разрывов) ДНК.
Разрывы эти в начале мейоза возникают не сами собой – их создает специальный фермент, эндонуклеаза SPO11.
Этот фермент является модификацией другого белка – ДНК-топоизомеразы VI, унаследованной эукариотами от своих прокариотических предков – архей.
Клетка начинает «чинить» разорванные молекулы ДНК при помощи древнего механизма гомологичной рекомбинации. А для этого нужно объединить попарно гомологичные молекулы ДНК (чтобы использовать неповрежденные участки одной молекулы в качестве «матрицы» для исправления повреждений в другой молекуле). На этом и основано попарное объединение хромосом в профазе I мейоза.
Белковые оси хромосом
Эукариотический геном значительно больше прокариотического и содержит больше «избыточной» ДНК (различных некодирующих последовательностей, мобильных генетических элементов и др.
) Большие размеры генома стимулировали «распадение» его на отдельные хромосомы (линейные, в отличие от единственной кольцевой хромосомы прокариот).
Как следствие, «оказалась полезной новация в структуре хромосом – белковые оси, к которым стали крепиться петли гигантской молекулы ДНК – эукариотической хромосомы».
В клетках эукариот, делящихся путем митоза, сестринские хроматиды (образовавшиеся в результате репликации исходной молекулы ДНК) соединены посредством белков-когезинов. Чтобы хроматиды могли разойтись к полюсам клетки (в анафазе митоза), когезины должны гидролизоваться и исчезнуть из межхроматидного пространства.
Один из когезинов (Rec8) «оказался подходящим субстратом для наслоения на него других мейоз-специфичных белков», из которых в начале мейоза строятся более сложные белковые оси, или «линейные элементы», к которым ДНК крепится петлями, приобретая вид «ершика».
Каждая петля содержит несколько тысяч пар нуклеотидов и крепится к линейному элементу только в одном месте, «кнопочно». Такая организация хромосомы позволяет хромосомным локусам легче находить гомологичные им локусы в хромосоме-партнере.
Так обстоит дело у некоторых низших эукариот, у которых нет «классического» мейоза, характерного для более высокоорганизованных форм.
Синаптонемные комплексы. Ароморфоз.
У эукариот с классическим мейозом «бывшие линейные элементы хромосом соединяются попарно с помощью белковой «застежки-молнии» в двухосевую структуру, именуемую синаптонемным комплексом (СК)». Это обеспечивает тесное соединение гомологичных хромосом. Ю.Ф.
Богданов считает появление СК важным ароморфозом. СК существует ограниченное время в течение профазы I мейоза и затем распадается. СК обнаружен у многих сотен видов – от одноклеточных водорослей, грибов и протистов до многоклеточных.
Предполагается несколько возможных функций СК:
1) Организация профазной мейотической хромосомы. Благодаря СК возникает билатеральная организация пары соединившихся гомологичных хромосом. Петли хроматина располагаются по обе стороны от СК.
Это позволяет точно сопоставить петли хроматина в трехмерном пространстве клеточного ядра.
«По-видимому, это простейший, если не единственный, способ внести порядок и точность в процесс взаимного узнавания локусов гомологичных хромосом и обеспечить достаточную точность (гомологию) рекомбинации в мейозе.»
2) СК не только соединяет гомологичные хромосомы, но и не дает им «склеиться», удерживая их на расстоянии 70-120 нм друг от друга.
В конце профазы I (стадия диплотены) СК распадается во всех локусах, кроме локусов хиазм (перекрещивания хроматид).
Гомологичные хромосомы, взаимно оттолкнувшиеся во всех локусах, кроме локусов хиазм, выстраиваются на экваторе веретена деления в метафазе I. Они готовы разойтись к полюсам, как только освободятся от хиазм.
3) СК необходим для формирования хиазм. СК также не позволяет хиазмам располагаться слишком близко друг от друга.
Иными словами, благодаря СК хроматиды могут «рваться» и обмениваться участками лишь в ограниченном количестве мест. Ю.Ф.
Богданов оценивает это как положительное явление, поскольку «чрезмерная частота кроссинговера – не столько «благо» комбинаторики, сколько угроза стабильности адаптационно выгодных фенотипов».
Происхождение белков СК
У разных групп эукариот СК строится по единому «плану», но при этом используются совершенно разные (структурно негомологичные) белки. Это похоже на постройку домов по сходному плану (стены, крыша, окна), но из совершенно разных материалов.
Материал не важен – «важно, чтобы СК выравнивал параллельно лежащие гомологичные хромосомы, сохранял между ними пространство, в котором происходит рекомбинация ДНК, и сохранял бы эти условия столь долго, сколько необходимо для завершения рекомбинации и формирования хиазм».
Сравнительный анализ структуры белков СК показал, что они возникли независимо у предков современных эукариот, давших начало разным крупным таксонам (растений, грибов, животных).
Ю.Ф.Богданов приводит убедительные доводы и экспериментальные факты, свидетельствующие о том, что структуры СК формируются путем самосборки из белковых молекул, подобно многим другим сложным молекулярным комплексам.
Автор отмечает, что «самоорганизующиеся структуры могут состоять из разных белковых субъединиц и нуклеиновых кислот, как это наблюдается в отношении вирусов и рибосом».
Важно, что белки, сходные по размерам и трехмерной организации, могут служить компонентами одинаковых структур у далеких друг от друга организмов даже в тех случаях, когда первичная структура (аминокислотная последовательность) этих белков сильно различается.
Почему в первом делении мейоза сестринские хроматиды не расходятся?
В отличие от митоза и второго деления мейоза, в первом делении мейоза сестринские хроматиды остаются «склееными», потому что ген CDC31, продукт которого нужен для гидролиза когезинов, в мейозе I оказывается временно отключен продуктом другого гена – SGO1 (у дрожжей S. cerevisae). Во время второго деления мейоза ген SGO1 не функционирует, «запрет» на работу CDC31, снимается, и сестринские хроматиды разделяются. У дрозофилы вместо SGO1