
Класс Круглоротые. Строение, размножение, разнообразие и значение Круглоротых. Отряды Миноги и Миксины. Надкласс Рыбы
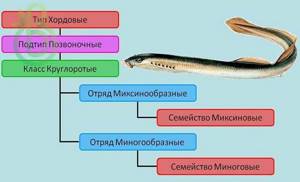
Тип Хордовые – Подтип Позвоночные – Класс Круглоротые: Отряд Миксинообразные – Семейство Миксиновые; Отряд Миногообразные – Семейство Миноговые.
Второе название Круглоротых — Бесчелюстные. Самые древние и примитивные представители позвоночных. Известно около 50 видов класса Круглоротые (Cyclostomata). Эти исключительно водные животные, которые ведут паразитический или полупаразитический образ жизни. Живут в пресных и соленых водоемах. К ним относят два отряда: Миноги и Миксины.
Строение Круглоротых: Миноги и Миксины
Внешнее строение Миноги: непарная ноздря, предротовая присасывательная воронка, глаза, наружные жаберные отверстия, спинные плавники, хвостовой плавник, мочеполовой сосочек и анальное отверстие
Миноги и Миксины имеют, удлиненное тело, округлое на поперечном разрезе. Главный и хвостовой отделы не отделяются. Нет парных конечностей.
Внешнее и внутреннее строение Миксины: усики, усик, редуцированные глаза, жаберные отверстия, глоточно-кожный проток, поры слизи, анальное отверстие, ноздря, рот, мозг, спинной мозг, хорда, язык с зубами, глотка, жаберный мешок и внешнее жаберное отверстие
Осевой скелет образован хордой. Вместе с нервной трубкой окружены соединительнотканной оболочкой. Хрящевой череп прикрывает головной мозг снизу и по бокам.
Сверху мозг закрыт соединительнотканной перепонкой (верхняя часть черепа еще не образована). Есть висцеральный череп, который образован хрящами и поддерживает ротовую воронку и язык.
Он включает висцеральный скелет жаберных мешков и околосердечный хрящ, окружающий сердце. Впереди к черепу примыкает обонятельная капсула, а по бокам — слуховые.
На голове расположена присасывающая воронка. На ее дне находится ротовое отверстие. Внутри воронки, а также на мышечном языке находятся роговые зубы. У миксин на месте ротовой воронки находятся короткие щупальца.
Мускулатура сегментированная. Мышцы туловища и хвоста состоят из отдельных миомеров, разделенных соединительнотканными перепонками (миосептами). В области жабр миомеры образуют спинную и брюшную мышечные ленты. Язык имеет собственную сложную мускулатуру. Напоминает мышечную систему ланцетника.
Покровы. Кожа голая, без чешуек. Состоит из двух слоев: эпидермиса и собственно кожи. В коже в эпидермисе много одноклеточных слизистых желез, поэтому тело густо покрыто слизью.
Пищеварительная система Круглоротых
Пищеварительная система Круглоротых подобна пищеварительной системе ланцетника. Начинается ртом. Глотка у миног функционирует только на личиночной стадии. У взрослых животных она разделена на дыхательную трубку и пищевод. Желудок не развит.
Пищевод переходит сразу в среднюю кишку. Кишки не имеют изгибов. Внутри кишки в слизистой оболочке формируется складка — спиральный клапан, который увеличивает всасывательную поверхность. Хорошо развита большая печень.
Задний конец кишечника наружу открывается анальным отверстием.
Язык действует как поршень и нагнетает кровь, жидкость в ротовую полость, а оттуда — в пищевод.
Органы выделения Круглоротых
Органы выделения Круглоротых представлены лентовидными парными почками. Они лежат по сторонам спинной аорты вдоль всей брюшной полости. Мочевыделительные каналы открываются общим с половой системой отверстием.
Кровеносная система Круглоротых
Кровеносная система Круглоротых: замкнутая у миног и незамкнутая у миксин. Один круг кровообращения. Сердце двухкамерное, состоит из предсердия и желудочка. Все отделы сердца содержат венозную кровь. Кровеносные сосуды делятся на артерии, вены и капилляры. Артерии несут кровь от сердца, вены — к сердцу.
От передней части желудочка отходит аорта, которая разветвляется на две выносные жаберные артерии. От сердца кровь по артериям поступает в жаберные мешки, а из них — во все органы. В жабрах происходит газообмен, кровь обогащается кислородом. В тканях кровь превращается в венозную. Венозная кровь собирается по венам в венозный синус.
В печени образуется ее воротная система. Кровь красная.
Органы дыхания Круглоротых
Сферические жаберные щели расположены по сторонам головы позади глаз и ведут к жаберным мешкам. Жаберные мешки имеют эктодермальное происхождение и развиваются в жаберных щелях.
В жаберных мешках много складок (жаберные лепестки), оплетенных капиллярами, в которых происходит газообмен. Вода всасывается через рот, выводится через жаберные отверстия наружу.
Во время питания минога всасывает и выводит воду через жаберные отверстия.
Нервная система Круглоротых
Нервная трубка разделена на спинной и головной мозг. Головной мозг имеет пять отделов: продолговатый мозг, мозжечок, средний мозг, промежуточный и передний мозг. Все отделы расположены в одной плоскости, то есть не образуют изгибов. От головного мозга отходят 10 пар головных нервов.
Органы чувств Круглоротых
По сторонам головы расположены глаза, прикрытые полупрозрачной кожей. Дополнительным светочувствительным органом является эпифиз, расположенный близко к коже головы.
Сверху на голове между глазами находится отверстие непарного органа обоняния (ноздря). По бокам на голове, вокруг и впереди глаз есть короткая боковая линия в виде очень мелких отверстий, которая является органом чувств и воспринимает колебания водной среды.
Орган слуха парный, представлен внутренним ухом (имеют два полукруглых канальца). В коже много разбросано клеток — механо-, термо- и хеморецепторов.
Размножение Круглоротых
Раздельнополые. Половые железы непарные. Они не имеют выводных протоков. Гаметы в полость тела попадают через разрывы стенок половых желез и выходят в воду через общие с выделительной системой отверстия. Оплодотворение внешнее. Развитие с метаморфозом у миног и прямое у миксин.
Разнообразие Круглоротых
Представители: речная минога, венгерская минога, украинская минога, миксина европейская и др.
Отряд Миноги (Petromyzoniformes)
Внешний вид Миноги: глаза, жабры, зубные пластинки, тело и хвост
Известно свыше 20 видов. Пресноводные и проходные животные. Общая продолжительность жизни — 5-6 лет. Большинство видов — эктопаразиты рыб.
Тело голое, покрытое слизью. Длина — 50-80 см (до 1 м). На спинной стороне — 1-2 кожных плавника. Хвостовой и спинной плавники поддерживаются рядом тонких хрящевых палочек. Хорошо развиты глаза. Полость тела выстелена тонкой эпителиальной оболочкой.
Миноги питаются, когда присасываются ротовой воронкой к рыбе. Зубцами вокруг рта и на языке пробуравливают кожу, вводят пищеварительные соки. Ткани перевариваются в жидкость, которую высасывает минога. Питаются кровью, выедают мышцы, внутренние органы.
С каждой стороны тела имеют по 7 жаберных щелей. У морских миног есть слабый электрический орган, который помогает ощущать приближение других организмов.
Для размножения заходят в небольшие пресные речки с быстрым течением. Икру откладывают в гнезда — ямки. Имеют большую плодовитость (от 0,8 тыс. до 200 тыс. икринок). Гибнут после размножения.
Развитие непрямое. Личинка миног — пескоройка. По строению похожа на ланцетника. Живет 2-4 года в норках на дне и питается мелкими беспозвоночными (по типу питания — фильтратор).
Потом превращаются во взрослых животных.
Минога речная (Lampetra fluviatilis)
Живет в речках. Длина тела — 40 см.
Парные конечности отсутствуют. На спинной стороне — два кожаных плавника. Последний из них переходит непосредственно в хвостовой плавник. Брюшная лопасть хвостового плавника сливается с небольшим анальным плавником. Сверху по краям хорды лежат небольшие хрящики — зачатки позвонков.
При питании присасывается к телу рыбы, прогрызает кожу и высасывает соки и ткани жертвы.
Отряд Миксины (Myxiniformes)
Внешний вид Миксины: кожа, тело, щупальца и рот
Живут исключительно в морях. Известно около 20 видов. Длина составляет от 50 см до 1 м. Устроены немного проще, чем миноги.
Тело очень гибкое. Не имеют спинного плавника. Жаберных щелей от 1 до 15. Рот не имеет губ. Он окружен мясистыми усиками. Усики — это орган осязания. Глаза недоразвитые, скрытые под кожей. Вдоль нижней поверхности тела расположены два ряда пор — отверстий подкожных слизистых желез.
По типу питания хищники или паразиты. Поедают икру, мертвых и живых рыб. Нападают на рыб, имеющих значительно большие размеры (даже на акул). Разрывают кожу жертвы с помощью зубцов языка и погружаются глубоко в ткани.
Миксины впрыскивают выделения слюнных желез в ткани рыбы, переваривают ее изнутри. Потом высасывают жидкость. Иногда от жертвы остается только кожа и кости.
Миксины, которые питаются мертвыми организмами, образуют ходы в их теле.
Кровеносная система незамкнутая. Сердце одно основное и три дополнительных.
Яйца откладывают летом (20-30 штук). Имеют округлую форму, сцеплены между собою. Прикрепляют их к водным предметам. Развитие прямое.
Значение Круглоротых
Миноги, как правило, присасываются к больным или слабым рыбам, поэтому являются естественными санитарами. Миног можно употреблять в пищу, они очень вкусные. Миксины промышленного значения не имеют.
Иногда миноги и миксины вредят рыбоводству и рыбной ловле.
Надкласс Рыбы
Надкласс Рыбы: Класс Хрящевые Рыбы (около 600 видов) и Класс Костные Рыбы (около 25 000 видов): Подкласс Кистеперые, Подкласс Двоякодышащие, Подкласс Лучеперые и Подкласс Хрящевые
К надклассу Рыбы (Pisces) принадлежат около 20 тыс. видов рыб, что составляет почти 50 % видов всех позвоночных. Живут исключительно в водной среде. Появляются парные конечности, челюсти. Дыхание происходит при помощи жабр. Тело большинства имеет обтекаемую форму и покрыто чешуей.
Рыбы имеют внутренний скелет (хрящевой или костный), состоящий из позвонков. У взрослых организмов хорда отсутствует.
Пищеварительный канал более дифференцирован — выделяется желудок, тонкий и толстый отделы кишечника. Прямая кишка открывается в клоаку.
Клоака — это полость, в которую открываются прямая кишка, мочеточники и половые протоки. Кровеносная система замкнутая, сердце двухкамерное. Один круг кровообращения. Большинство рыб имеет хорошо развитые органы чувств.
Носовые ходы парные. Типичным органом чувств для рыб является боковая линия.
Надкласс делят на два класса: Хрящевые и Костные рыбы. Преобладающее количество рыб относится к Костным.
Подцарство многоклеточные
Нервная система и органы чувств
Ощущение электрического поля — электрорецепция — присущее многим видам рыб — не только тем, которые могут сами генерировать электрические разряды.
Электрические сигналы улавливаются с помощью специальных ямками на поверхности тела.
Эти ямки заполнены гелеподобным веществом, которое проводит электрический ток и содержит в себе электрорецепторные клетки, которые образовывают синапсы с нейронами.
Спинной мозг проходит внутри нервных дуг позвонков по всей длине позвоночника рыбы. Аналогично миомерам и позвоночнику, в строении спинного мозу наблюдается сегментация. В каждом сегменте тела сенсорные нейроны входят в спинной мозг через дорсальные корешки, а двигательные нейроны выходят из него через вентральные.
Глаза рыб по своему строению очень похожи с глазами других позвоночных.
Главное принципиальное отличие рыбьего глазу заключается в том, что для фокусирования на предмете, рыбы не изменяют кривизну хрусталика, а приближают или отдаляют его от роговицы.
Структура сетчатки варьирует для рыб в зависимости от места их обитания: у глубоководных видов глаза приспособлены для восприятия света преимущественно красной части спектра, а рыбы, которые живут на мелководье, воспринимают более широкий спектр.
Нюх и вкус позволяют рыбам ориентироваться в химическом составе окружающей среды.
Способность рыб к ощущению химических сигналов хорошо иллюстрируется лососями, которые идя на нерест из моря к речным системам, определяют по вкусу воды именно тот ручей, в которой когда-то самые вышли из икры.
Обонятельные рецепторы рыб расположены в ноздрях (которые, в отличие от других позвоночных, не соединяются с носоглоткой), а вкуса — не только в ротовой пустоте, но часто и на жаберных структурах, усиках, и даже на плавниках и на коже.
Главными механорецепторами рыб являются органы слуха, которые функционируют как орган и слуха, и равновесия, а также органы боковой линии.
Внутреннее ухо пластинчастожабернsх (акул и скатов) и костистых рыб состоит из трех полукруглых каналов, расположенных в трех взаимно-перпендикулярных плоскостях, и трех камер, каждая из которых вмещает отолиты.
Некоторые виды рыб (например, серебряный карась и разные виды сомов) имеют комплекс косточек, которые называются Вебберов аппарат и соединяют ухо с плавательным пузырем. Благодаря этой адаптации внешние вибрации усиливаются плавательным пузырем, как резонатором. Отолит в третьей камере, обеспечивает рыбе ориентацию в пространстве.
Перемещение воды по поверхности рыбы ощущается структурами, которые называются нейромасты. Эти органы могут быть рассеяны поодиночке, или собраны под чещуей в совокупности каналов, которые называются боковой линией.
Нейромасты включают гелевой консистенции полушарие (капулу) и сенсорные волосовидные клетки, а также синапсы нервных волокон, которые образовываются на волосовидних клетках. Перемещение воды служит причиной отгиба волосовидных клеток, который превращается в нервные импульсы.
Эти импульсы позволяют составить довольно подробную картину окружающей среды: некоторые виды рыб, лишенные глаз, целиком ориентируются и перемещаются, полагаясь только на органы боковой линии.
Ощущение электрического поля — электрорецепция — присущее многим видам рыб — не только тем, которые могут сами генерировать электрические разряды.
Электрические сигналы улавливаются с помощью специальных ямками на поверхности тела.
Эти ямки заполнены гелеподобным веществом, которое проводит электрический ток и содержит в себе электрорецепторные клетки, которые образовывают синапсы с нейронами.
Обновлено (10.12.2010 22:49)
Нервная система и органы чувств [1974 Вавилкин А.С., Иванов А.П., Куранова И.И. – Основы ихтиологии и рыбоводства]
Нервная система рыбы, как и других животных, обеспечивает согласованную деятельность всех органов и связывает организм с окружающей средой. Нервная система разделяется на центральную – головной и спинной мозг и периферическую – головные и спинные нервы. Нервы головного и спинного мозга в виде белых нитей подходят ко всем органам и мышцам. Различают нервы чувствительные и двигательные. По чувствительным нервам возбуждение проводится от органов чувств и других органов к мозгу, по двигательным нервам возбуждение передается в обратном направлении, т. е. от мозга к органам. Вся центральная нервная система, т. е. головной и спинной мозг, образована телами нервных клеток с их отростками, а периферическая – нервами, отходящими от центральной нервной системы. Нерв – это несколько отростков нервной клетки, одетых в оболочку. Специфической особенностью элементов нервной системы является их способность воспринимать раздражение извне, Ответная реакция организма на раздражение, осуществляемая нервной системой, носит название рефлекса. При всех рефлексах происходит раздражение окончаний чувствующих нервов-рецепторов. Возбуждение, возникшее в рецепторах, по чувствующему волокну передается в центральную нервную систему, оттуда по двигательным нервам поступает к мышцам тех или других органов, вызывая их деятельность. Путь, по которому идет возбуждение, называется рефлекторной дугой.
Головной мозг рыб довольно примитивен, имеет линейное расположение своих частей. Он состоит из пяти основных отделов: передний мозг (у высших животных называемый полушариями большого мозга), промежуточный мозг, средний мозг, мозжечок и продолговатый мозг.
Продолговатый мозг переходит в спинной, расположенный в верхних дугах позвоночника (рис. 18). Промежуточный мозг имеет вырост, называемый гипофизом. Гипофиз – железа внутренней секреции.
Впрыскивание созревающим самкам рыб экстракта гипофиза чрезвычайно ускоряет созревание икры и применяется с этой целью в промышленном рыбоводстве. Передний мозг является центром обоняния.
Рис. 18. Строение головного мозга окуня: 1 – носовая капсула; 2 – обонятельные доли; 3 – передний мозг; 4 – средний мозг; 5 – мозжечок; 6 – продолговатый мозг; 7 – спинной мозг; 8 – глазничная ветвь; 9 – слуховой нерв; 10 – блуждающий нерв
Промежуточный мозг выполняет функции переключателя возбуждений, идущих из всех частей мозга, связанных с ним. Средний мозг является центром зрительных нервов. У костистых Рыб он сильно развит и состоит из массивного основания и крышки, которая разделена на два полушария – зрительные доли. Мозжечок является регулирующим аппаратом для координации движения.
У костистых рыб он хорошо развит, особенно у быстро плавающих. Продолговатый мозг соединяет различные центры со спинным мозгом. В нем залегает наиболее важный жизненный центр – дыхательный. Центр слуха и осязания также находится в продолговатом мозгу.
Относительная величина этих отделов соответствует роли различных чувств в жизни рыбы, и внешний вид мозга позволяет судить об образе жизни рыбы.
От головного мозга отходят 10 пар нервов: обонятельный, зрительный, глазодвигательный, блоковый, тройничный, отводящий, лицевой, слуховой, языкоглоточный и блуждающий.
Спинной мозг рыб представляет собой шнур цилиндрической формы, проходящий в верхних дугах позвоночника. От него отходит много спинномозговых нервов, число которых соответствует количеству позвонков.
Симпатическая нервная система у рыб состоит из скоплений нервных клеток, или ганглиев, лежащих по бокам позвоночника. Функциональное значение симпатической системы – иннервация гладких мышц внутренних органов. Деятельность симпатической нервной системы и гладких мышц непроизвольна.
Рыбы
Конечный мозг (Telencephalon) хрящевых и костистых рыб принципиально отличаются друг от друга по структуре, развитию и уровню функциональной организации, поэтому их необходимо разбирать отдельно.
Конечный мозг (большие полушария) хрящевых рыб построен по инвертированному типу, т.е.
содержат латеральные мозговые желудочки, хотя степень их развития в разных группах акул и скатов различается.
В полушариях акул и скатов выделяют две основные области: дорсальную (паллиальную) и вентральную (субпаллиальную), гомологичных соответствующим областям мозга наземных позвоночных (рис. 11).
Рис. 11. Структура полушарий конечного мозга акулы Squalus acanthyas L. (по: Обухов, 1999, с изменениями).
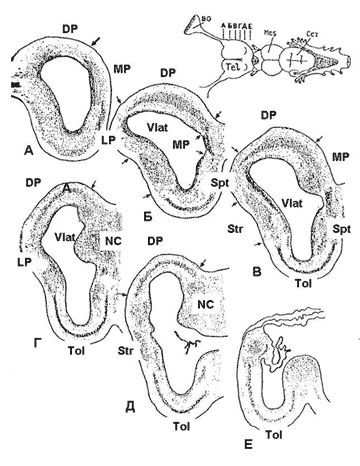 Рис. 11. Структура полушарий конечного мозга акулы Squalus acanthyas L. (по: Обухов, 1999, с изменениями).
Рис. 11. Структура полушарий конечного мозга акулы Squalus acanthyas L. (по: Обухов, 1999, с изменениями). А-Е – уровни срезов мозга, DP, MP, LP – дорсальный, медиальный и латеральный паллиум, Str – стриатум (субпаллиум), Spt – септум, Tol – обонятельный бугорок, Vlat – боковой желудочек (полость конечного мозга), NС – центральное ядро, стрелки – границы участков полушария.
Следует отметить, что у ряда видов акул в дорсальном паллиуме формируется настоящий корковый слой с дифференцированными формами нейронов, сходными с таковыми в неокортексе млекопитающих и птиц!
Высокий уровень цитоархитектонической и нейронной дифференцировки ряда отделов конечного мозга хрящевых рыб коррелирует с высоким уровнем функциональной организации конечного мозга у отдельных видов акул и скатов.
В полушария их головного мозга проецируются все основные сенсорные системы, а биохимический состав нейрональных мембран сходен с таковым у млекопитающих. Система афферентных и эфферентных проекций также аналогичны.
Формирование у ряда хрящевых рыб в ЦНС структур, сходных по уровню морфо-функциональной организации с высшими позвоночными животными – есть проявление эволюционного феномена «филогенетического опережения».
Это явление связано с появлением в ходе эволюции у отдельных представителей определенной группы животных признаков, выходящих за рамки «типовых» характеристик группы, что отражает мозаичный характер эволюции. В целом направление, в котором развивалась ЦНС ряда видов хрящевых рыб, можно рассматривать как эволюционно-прогрессивное (Обухов, 1999).
Среди костных рыб Лучеперые рыбы являются единственной группой позвоночных животных, чей конечный мозг построен по эвертированному типу (рис. 12).
Процессу эверсии подвержена дорсальная часть полушария. По одной из классификаций она разделяется на ряд цитоархитектонических зон: медиальную, дорсальную, дорсо-латеральную, латеральную и центральную. Вентральная (субпаллиальная) область также подразделена на ряд зон.
В связи с особенностями эмбриогенеза конечного мозга лучеперых рыб сложным является вопрос гомологии их полушарий со структурами конечного мозга инвертированного типа.
Одни исследователи предлагают проводить прямую гомологию зон мозга двух типов, другие считают невозможным проведение каких-либо сравнений, поскольку рассматривают конечный мозг лучеперых рыб как особую линию в эволюции мозга позвоночных.
Третьи, а их большинство, основываясь на современных данных о морфологии, гистохимии и связях конечного мозга позвоночных и учитывая особенности процессов инверсии и эверсии, считают, что можно найти решение данной проблемы.
Так, зона Dp рассматривается как возможный гомолог латерального паллиума, зоны Dl и Dd – медиального и дорсального паллиума наземных позвоночных животных соответственно. Каудальные отделы зоны Dm и часть зоны Dc сравнивают со стриатумом, отмечая при этом, что другие части этих зон включают компоненты дорсального паллиума. Ростральные отделы медиальной зоны гомологизируются с частью амигдалы. Вентральные зоны Vd и Vv сравниваются с областью септальных ядер, зона Vl – с обонятельным бугорком, а каудальные отделы Vs, Vp, Vi – с частью амигдалы.
Среди огромного количества видов и групп Лучеперых рыб встречается разный уровень дифференцировки полушарий: от наиболее просто устроенного полушария у многоперых и осетровых рыб, до более сложной структуры полушарий у ряда высших костистых рыб. Отмечается четкая эволюционная линия развития конечного мозга Лучеперых рыб: Chondrostei – Holostei – Teleostei (рис. 13).
Рис. 13. Схемы цитоархитектонического строения полушарий ряда видов Лучеперых рыб (А) и фрагмент нейронной структуры конечного мозга карпа (Б) (по: Обухов, 1975; Никоноров, Обухов, 1983).
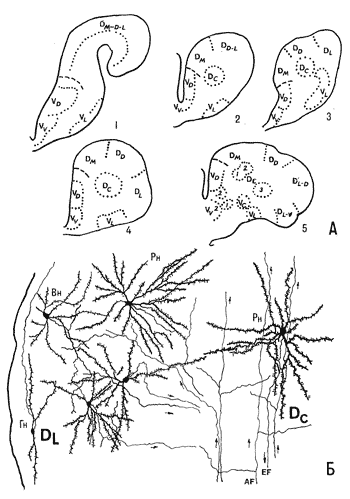 Рис. 13. Схемы цитоархитектонического строения полушарий ряда видов Лучеперых рыб (А) и фрагмент нейронной структуры конечного мозга карпа (Б) (по: Обухов, 1975; Никоноров, Обухов, 1983).
Рис. 13. Схемы цитоархитектонического строения полушарий ряда видов Лучеперых рыб (А) и фрагмент нейронной структуры конечного мозга карпа (Б) (по: Обухов, 1975; Никоноров, Обухов, 1983). 1 – Polypterus palmas L., 2 – Aciperser baeri Br., 3 – Lepomis punctatus, 4 – Salmo salar L., 5 – Ictalurus punctatus L. DL, DC – зоны дорсальной области полушария карпа Ciprinus carpio L., вн, гн, рн – веретеновидные, горизонтальные и радиальные нейроны, af, ef – афферентные и эфферентные волокна.
Анализ нейронной структуры полушарий конечного мозга различных представителей лучеперых рыб показал, что общий уровень его организации достаточно низок.
Основная масса нейронов паллиума представлена разновидностями одного – аллодендритного класса нейронов.
Это мультиполярные радиальные нейроны, расположенные преимущественно в глубоких слоях периферических зон паллиума и в центральной зоне, а также веерообразные и горизонтальные нейроны, сосредоточенные в поверхностных слоях зон (рис. 13 и рис. 14).
Степень ветвления дендритов не превышает 3-4-х классов, аксон часто отходит не от тела нейрона, а от начального сегмента одного из дендритных стволов.
Клетки сочетают в себе признаки ассоциативных и проекционных элементов, поскольку некоторые из их аксонных коллатералей ветвятся в пределах данной зоны, а другие – длинные проекционные ветви выходят за пределы зоны и даже области полушария.
У большинства же других позвоночных (как высших, так и низших) популяции нейронов паллиума разделены на субпопуляции ассоциативных интернейронов и проекционных нейронов.
В вентральной, субпаллиальной области полушарий нейронная дифференцировка еще слабее – здесь преобладают нейроны лептодендритного типа. У лучеперых рыб в паллиуме и субпаллиуме не обнаружено высокодифференцированных короткоаксонных звездчатых нейронов, являющихся критерием высокого уровня организации нервных центров.
В целом в эволюционном ряду лучеперых рыб (Chondrostei – Holostei – Teleostei) не происходит существенных изменений в уровне нейронной дифференцировки полушарий, что позволило выдвинуть гипотезу об особом, эволюционно-консервативном пути развития головного мозга Лучеперых рыб. Это подтверждается данными по физиологии, системе связей и поведении этих рыб.
Нервная система и органы чувств
Нервная система рыб представлена центральной нервной системой и связанной с ней периферической и вегетативной (симпатической) нервной системой. Центральная нервная система состоит из головного и спинного мозга.
К периферической нервной системе относятся нервы, отходящие от головного и спинного мозга к органам. Вегетативная нервная система в основе имеет многочисленные ганглии и нервы, иннервирующие, мышцы внутренних органов и кровеносных сосудов сердца.
Нервная система рыб по сравнению с нервной системой высших позвоночных характеризуется рядом примитивных черт.
Центральная нервная система имеет вид нервной трубки, тянущейся вдоль туловища; часть ее, лежащая над позвоночником и защищенная верхними дугами позвонков, образует спинной мозг, а расширенная передняя часть, окруженная хрящевым или костным черепом, составляет головной мозг.
Трубка имеет внутри полость (невроцель), представленную в головном мозгу желудочками мозга.
В толще мозга различают серое вещество, слагающееся из тел нервных клеток и коротких отростков (дендритов), и белое вещество, образованное длинными отростками нервных клеток – нейритами или аксонами.
Общая масса мозга у рыб мала: она составляет в среднем у современных хрящевых рыб 0,06 – 0,44%, у костных – 0,02 – 0,94% от массы тела, в том числе у налима 1/700 массы тела, щуки 1/3000, акулы – 1/37000 , в то время как у летающих птиц и млекопитающих 0,2 – 8,0 и 6,3 – 3,0%.
В строении головного мозга сохраняются примитивные черты: отделы мозга располагаются линейно. В нем выделяют передний мозг, промежуточный, средний, мозжечок и продолговатый, переходящий в спинной мозг.
Полости переднего, промежуточного и продолговатого мозга называются желудочками: полость среднего мозга–сильвиевым водопроводом (она соединяет полости промежуточного и продолговатого мозга, т. Е.
третий и четвертый желудочки).
Передний мозг благодаря продольной борозде имеет вид двух полушарий. К ним прилегают обонятельные луковицы (первичный обонятельный центр) или непосредственно (у большинства видов), или через обонятельный тракт (карповые, сомовые, тресковые).
В крыше переднего мозга нет нервных клеток. Серое вещество в виде полосатых тел сосредоточено главным образом в основании и обонятельных долях, выстилает полость желудочков и составляет главную массу переднего мозга. Волокна обонятельного нерва связывают луковицу с клетками обонятельной капсулы.
Передний мозг является центром обработки информации, поступающей от органов обоняния. Благодаря своей связи с промежуточным и средним мозгом он участвует в регуляции движения и поведения. В частности, передний мозг принимает участие в формировании способности к таким актам, как икрометание, охрана икры, образование стаи и т. Д.
В промежуточном мозге развиты зрительные бугры. От них отходят зрительные нервы, образующие хиазму (перекрест, т. Е. часть волокон правого нерва переходит в левый нерв и наоборот). На нижней стороне промежуточного мозга (гипоталамус) имеется воронка, к которой прилегает гипофиз; в верхней части промежуточного мозга развивается эпифиз. Гипофиз и эпифиз являются железами внутренней секреции.
Промежуточный мозг выполняет многочисленные функции. Он воспринимает раздражения от сетчатки глаза, участвует в координации движений, в переработке информации от других органов чувств. Гипофиз и эпифиз осуществляют гормональную регуляцию обменных процессов. Средний мозг наибольший по объёму.
Он имеет вид двух полушарий (зрительные доли). Зрительные доли являются первичными зрительными центрами, воспринимающими возбуждение. Из этих долей берут начало волокна зрительного нерва.
В среднем мозгу обрабатываются сигналы, идущие от органов зрения и равновесия; здесь помещаются центры связи с мозжечком, продолговатым и спинным мозгом.
Мозжечок расположен в задней части мозга и может иметь форму или маленького бугорка, прилегающего сзади к среднему мозгу, или большого мешковидно-вытянутого образования, примыкающего сверху к продолговатому мозгу.
Особенно большого развития достигает мозжечок у сомов, а у мормируса относительная его величина является наибольшей среди остальных позвоночных. В мозжечке рыб, как и высших позвоночных, имеются клетки Пуркинье. Мозжечок является центром всех моторных иннервации при плавании, схватывании пищи.
Он обеспечивает координацию движений, поддержание равновесия, мышечную деятельность, связан с рецепторами органов боковой линии.
Пятый отдел головного мозга, продолговатый мозг, без резкой границы переходит в спинной мозг. Полость продолговатого мозга –четвертый желудочек – продолжается в полость спинного мозга – невроцель. Значительная масса продолговатого мозга состоит из белого вещества.
От продолговатого мозга отходит большая часть (шесть из десяти) черепно-мозговых нервов. Он является центром регуляции деятельности спинного мозга и вегетативной нервной системы.
В нем располагаются наиболее важные жизненные центры, регулирующие деятельность дыхательной, скелетно-мышечной, кровеносной, пищеварительной, выделительной систем, органов слуха и равновесия, вкуса, боковой линии, электрических органов у имеющих их рыб и т. Д.
Поэтому при разрушении продолговатого мозга, например при перерезке туловища позади головы, наступает быстрая смерть рыбы. Через приходящие в продолговатый мозг спинномозговые волокна осуществляется связь продолговатого и спинного мозга.
Передний мозг (и обонятельные доли) относительно сильнее развит у хрящевых рыб (акулы и скаты) и слабее – у костистых. У малоподвижных, например донных, рыб мозжечок мал, но сильнее развиты передний и продолговатый отделы мозга в соответствии с большой ролью обоняния и осязания в их жизни (камбалы).
У хорошо плавающих рыб (пелагических, питающихся планктоном, или хищничающих), наоборот, гораздо большее развитие получают средний мозг (зрительные доли) и мозжечок (в связи с необходимостью быстрой координации движения). Рыбы, обитающие в мутной воде, имеют маленькие зрительные доли, небольшой мозжечок.
Слабо развиты зрительные доли у глубоководных и слепых рыб.
Спинной мозг является продолжением продолговатого мозга. Он имеет форму округлого тяжа и лежит в канале, образованном верхними дугами позвонков. В спинном мозге серое вещество расположено внутри, а белое–снаружи.
От спинного мозга метамерно, соответственно каждому позвонку, отходят спинномозговые нервы, иннервирующие поверхность тела, туловищные мышцы, а благодаря соединению спинномозговых нервов с ганглиями симпатической нервной системы – и внутренние органы.
У костистых рыб ганглии вегетативной нервной системы соединяются двумя продольными нервными стволами. Соединительные ветви ганглиев связывают вегетативную нервную систему с центральной. Взаимосвязи центральной и вегетативной нервной систем создают возможность некоторой взаимозаменяемости нервных центров.
Вегетативная нервная система действует в определённой степени автономно, независимо от центральной нервной системы и определяет непроизвольную, автоматическую деятельность внутренних органов даже в том случае, если ее связь с центральной нервной системой нарушена.
Реакцию организма рыбы на внешние и внутренние раздражения определяет рефлекс. У рыб можно выработать условный рефлекс на свет, форму, запах, вкус, звук. По сравнению с высшими позвоночными у рыб условные рефлексы образуются медленнее, а гаснут быстрее.
Тем не менее и аквариумные, и прудовые рыбы вскоре после начала регулярного кормления скапливаются в определённое время у кормушек.
Привыкают они и к звукам во время кормления (постукивание по стенкам аквариума, звон колокольчика, свист, удары) и какое-то время подплывают на эти раздражители и при отсутствии пищи.
Органы зрения рыб устроены в основном так же, как у других позвоночных.
Сходен с остальными позвоночными у них и механизм восприятия зрительных ощущений: свет проходит в глаз через прозрачную роговицу, далее зрачок – отверстие в радужной оболочке – пропускает его на хрусталик, а хрусталик передает фокусирует свет на внутреннюю стенку глаза сетчатку, где и происходит его непосредственное восприятие. Сетчатка состоит из светочувствительных (фоторецепторные), нервных, а также опорных клеток.
Светочувствительные клетки располагаются со стороны пигментной оболочки. В их отростках, имеющих форму палочек и колбочек, имеется светочувствительный пигмент.
Количество этих фоторецепторных клеток очень велико –на 1 мм2 сетчатки у карпа их насчитывается 50 тыс. (у кальмара –162 тыс. , паука–16 тыс. , человека – 400 тыс. , совы – 680 тыс.).
Посредством сложной системы контактов конечных разветвлений чувствующих клеток и дендритов нервных клеток световые раздражения поступают в зрительный нерв.
Колбочки при ярком свете воспринимают детали предметов и цвет. Палочки воспринимают слабый свет, но детального изображения создать не могут.
Положение и взаимодействие клеток пигментной оболочки, палочек и колбочек меняются в зависимости от освещенности.
На свету пигментные клетки расширяются и прикрывают находящиеся около них палочки; колбочки подтягиваются к ядрам клеток и таким образом передвигаются к свету.
В темноте к ядрам подтягиваются палочки (и оказываются ближе к поверхности); колбочки приближаются к пигментному слою, а сократившиеся в темноте пигментные клетки прикрывают их.
Количество рецепторов разного рода зависит от образа жизни рыб. У дневных рыб в сетчатке превалируют колбочки, у сумеречных и ночных – палочки: у налима палочек в 14 раз больше, чем у щуки.
У глубоководных рыб, живущих в темноте глубин, колбочек нет, а палочки становятся больше и количество их резко увеличивается – до 25 млн/мм2 сетчатки; вероятность улавливания даже слабого света возрастает.
Большая часть рыб различает цвета, что подтверждается возможностью выработки у них условных рефлексов на определённый цвет – синий, зеленый, красный, жёлтый, голубой.
Некоторые отступления от общей схемы строения глаза рыбы связаны с особенностями жизни в воде. Глаз рыбы эллипсовидный. В числе других он имеет серебристую оболочку (между сосудистой и белковой), богатую кристалликами гуанина, которая придает глазу зеленовато-золотистый блеск.
Роговица почти плоская (а не выпуклая), хрусталик шаровидный (а не двояковыпуклый) – это расширяет поле зрения.
Отверстие в радужной оболочке – зрачок – может изменять диаметр только в небольших пределах. Век у рыб, как правило, нет.
Лишь акулы имеют мигательную перепонку, закрывающую глаз как занавеска, и некоторые сельди и кефали – жировое веко – прозрачную пленку, закрывающую часть глаза.
Расположение глаз по бокам головы (у большинства видов) является причиной того, что рыбы обладают в основном монокулярным зрением, а способность к бинокулярному зрению весьма ограничена.
Шаровидность хрусталика и перемещение его вперед к роговице обеспечивает широту поля зрения: свет в глаз попадает со всех сторон. Угол зрения по вертикали составляет 150°, по горизонтали– 168–170°. Но вместе с тем шаровидность хрусталика обусловливает близорукость рыб.
Дальность их зрения ограничена и колеблется в связи с мутностью воды от нескольких сантиметров до нескольких десятков метров.
Видение на дальние расстояния становится возможным благодаря тому, что хрусталик может быть оттянут специальной мышцей–серповидным отростком, идущим от сосудистой оболочки дна глазного бокала.
При помощи зрения рыбы ориентируются и относительно предметов, находящихся на земле.
Улучшение зрения в темноте достигается наличием отражательного слоя (тапетум) – кристалликов гуанина, подстилаемых пигментом.
Этот слой не пропускает свет к лежащим позади сетчатки тканям, а отражает его и возвращает вторично на сетчатку. Так увеличивается возможность рецепторов использовать свет, попавший в глаз.
В связи с условиями обитания глаза рыб могут сильно видоизменяться. У пещерных или абиссальных (глубоководных) форм глаза могут редуцироваться и даже исчезать.
Некоторые же глубоководные рыбы, наоборот, имеют огромные глаза, позволяющие улавливать совсем слабые следы света, или телескопические глаза, собирающие линзы которых рыба может поставить параллельно и обрести бинокулярное зрение.
Глаза некоторых угрей и личинок ряда тропических рыб вынесены вперед на длинных выростах (стебельчатые глаза).
Роль зрения как источника информации из внешнего мира для большинства рыб очень велика: при ориентации во время движения, при отыскивании и захвате пищи, при сохранении стаи, в нерестовый период (восприятие оборонительных и агрессивных поз и движений самцами-соперниками, а между особями разных полов – брачного наряда и нерестового «церемониала»), в отношениях жертва –хищник и т. Д.
Известно, что рыбы разных видов неодинаково реагируют на свет разной интенсивности и разной длины волны, т. Е. разного цвета. Так, яркий искусственный свет привлекает одних рыб (каспийская килька, сайра, ставрида, скумбрия и др.
) и отпугивает других (кефаль, минога, угорь и т. Д. ). Так же избирательно относятся разные виды к разным цветам и разным источникам света – надводным и подводным.
Все это положено в основу организации промышленного лова рыбы на электросвет (так ловят кильку, сайру и других рыб).
§ 34. Нервная система, поведение и размножение рыб
Нервная система рыб состоит из центральной и периферической. Центральная нервная система включает головной и спинной мозг. Головной мозг состоит из пяти отделов: передний, промежуточный, средний, мозжечок и продолговатый. Периферическая нервная система представлена нервами, идущими ко всем органам и мускулатуре.
Нервная система ланцетника представлена нервной трубкой, состоящей из нервных клеток и тянущейся вдоль всего тела над хордой. Головного мозга нет. От нервной трубки отходят многочисленные нервы к внутренним органам и поверхности тела.
Органы чувств позволяют рыбам хорошо ориентироваться в окружающей среде. Орган зрения — глаза, которые располагаются в глубоких впадинах черепа. Рыбы видят на сравнительно близких расстояниях и различают только форму и цвет предмета.
Органы обоняния — парные обонятельные мешки, выстланы чувствительными клетками. Они сообщаются с внешней средой через ноздри.
Орган слуха и равновесия представлен одним отделом — внутренним ухом, расположенным в задней части черепа и не имеющим связи с внешней средой. Звуковые волны передаются через кости черепа непосредственно внутреннему уху.
Орган вкуса представлен чувствительными клетками, которые расположены не только в ротовой полости, но и по всей поверхности тела, как и осязательные клетки. У некоторых рыб на голове есть осязательные усики (сазан, сом, треска).
Боковая линия — специальный орган рыб, воспринимающий давление воды. Это канал в толще кожи по бокам тела. Он сообщается с внешней средой рядом отверстий и содержит чувствительные клетки.
Эти клетки воспринимают направление и силу тока воды, а также очень тонкие звуковые колебания.
Благодаря боковой линии рыбы могут обходить препятствия, не видя их, ориентироваться в воде и держаться в стае.
В поведении рыб проявляются безусловные (врождённые) и условные (приобретённые) рефлексы. Безусловные рефлексы передаются по наследству. Условные рефлексы не передаются по наследству и вырабатываются в течение всей жизни. Основными среди безусловных являются оборонительный и пищевой рефлексы.
Если попытаться коснуться тела рыбы, она моментально метнётся в сторону. В этом случае мы наблюдаем оборонительный рефлекс. Если кормление сопровождать какими-либо действиями (условиями), например зажиганием лампочки или постукиванием по стеклу, то через некоторое время такой сигнал начинает привлекать рыб сам по себе, без подкормки.
На подобные сигналы у рыб вырабатываются приобретённые, или условные, рефлексы.
Сложное инстинктивное поведение рыб в период размножения называют нерестом.
Чем большее количество икринок, тем выше вероятность, что хоть какая-то часть из них вырастет во взрослую рыбу и та сможет продлить потомство. Много хищников обитает в водоемах, питающихся как икрой, так и мальками, которые очень уязвимы. Кроме того, на развитие негативно влияют и другие факторы (браконьерство, химические загрязнения).
Самки и самцы перемещаются в места водоёмов, более благоприятные для развития их потомства.
Рыбы одних видов устремляются из моря в реки (например, лососевые рыбы: кета, горбуша), другие — наоборот, идут из рек в море (речной угорь). Это так называемые проходные рыбы.
Для икрометания они проходят большие расстояния. Рыбы, живущие и размножающиеся в водоёме одного типа (карась, окунь, щука), называются оседлыми рыбами.
Рыба Тиляпия – вынашивает икру во рту и мальков, в случае опасности подает сигнал, также прячет во рту. Тихоокеанские и атлантические лососи хвостом вырывают в грунте гнезда откладывают в них икру, оплодотворяют ее и засыпают гравием. Самец Колюшки строит гнездо из растительных остатков в виде муфточки и охраняет икру.
Самец Судака расчищает на дне место для будущей кладки икры, затем охраняет ее, очищает от ила, смывая его сильными движениями грудных плавников. Если кладка остается без сторожевого самца, то охрану продолжает другой. Лабиринтовые рыбы строят гнездо из пузырьков воздуха, обволакивая их клейкими выделениями изо рта.
Самец Пинагора охраняет кладку икры, отложенную в литоральной зоне и при обсыхании поливает икру водой изо рта.
Рыбы, которые оберегают икру (например, сом), откладывают ее немного.
Причем у одного вида рыбы икра созревает полностью и нерест продолжается в течение короткого времени, в других она созревает и откладывается отдельными порциями, а перерывы между ее откладыванием длятся от нескольких дней до нескольких недель.
Таких нерестов у некоторых рыб бывает до нескольких десятков. Разовый нерест характерный для щуки, окуня, осетра. Порционный нерест — для карпа, густеры, карася, линя, которые откладывают икру за 2-3 приема.






