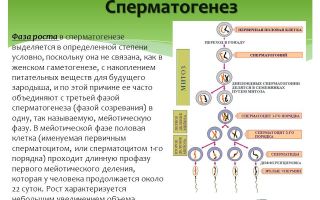
Сперматогенез, фазы и превращение клеток. Биологическое значение полового размножения
Гаметогенез(от гаметы и греч. genesis — происхождение), процесс развития и формирования половых клеток — гамет.Гаметы (от греч.
γᾰμετή — жена, γᾰμέτης — муж) — репродуктивные клетки, имеющие гаплоидный (одинарный) набор хромосом и участвующие в половом размножении.
При слиянии двух гамет в половом процессе образуется зигота, развивающаяся в особь (или группу особей) с наследственными признаками обоих родительских организмов.
Г. мужских гамет (сперматозоидов, спермиев) называют сперматогенезом, женских гамет (яйцеклеток) — овогенезом.
●У многоклеточных животных Г. происходит в специальных органах — половых железах, или гонадах (яичниках, семенниках, гермафродитных половых железах), и складывается из трёх основных этапов:
1) размножениепервичных половых клеток — гаметогониев (сперматогониев и оогониев) путём ряда последовательных митозов,
2) рост и созревание этих клеток, называют теперь гаметоцитами (сперматоцитами и ооцитами), которые, как и гаметогонии, обладают полным (большей частью диплоидным) набором хромосом.
В это время совершается основное событие Г.
у животных — деление гаметоцитов путём мейоза, приводящее к редукции (уменьшению вдвое) числа хромосом в этих клетках и превращению их в гаплоидные клетки
3) формирование сперматозоидов (либо спермиев) и яйцеклеток; при этом яйцеклетки одеваются рядом зародышевых оболочек, а сперматозоиды приобретают жгутики, обеспечивающие их подвижность. У самок многих видов животных мейоз и формирование яйца завершаются после проникновения сперматозоида в цитоплазму ооцита, но до слияния ядер сперматозоида и яйцеклетки.
●У растений Г. отделен от мейоза и начинается в гаплоидных клетках — в спорах (у высших растений — микроспоры и мегаспоры).
Из спор развивается половое поколение растения — гаплоидный гаметофит, в половых органах которого — гаметангиях (мужских — антеридиях, женских — архегониях) путём митозов происходит Г.
Исключение составляют голосеменные и покрытосеменные растения, у которых сперматогенез идёт непосредственно в прорастающей микроспоре — пыльцевой клетке. У всех низших и высших споровых растений Г. в антеридиях — это многократное деление клеток, в результате которого образуется большое число мелких подвижных сперматозоидов.
Г. в архегониях — формирование одной, двух или нескольких яйцеклеток. У голосеменных и покрытосеменных растений мужской Г. состоит из деления (путём митоза) ядра пыльцевой клетки на генеративное и вегетативное и дальнейшего деления (также путём митоза) генеративного ядра на два спермия. Это деление происходит в прорастающей пыльцевой трубке.
Женский Г. у покрытосеменных растений — обособление путём митоза одной яйцеклетки внутри 8-ядерного зародышевого мешка. Основное различие Г. у животных и растений: у животных он совмещает в себе превращение клеток из диплоидных в гаплоидные и формирование гаплоидных гамет; у растений Г. сводится к формированию гамет из гаплоидных клеток.
СПЕРМАТОГЕНЕЗ– это развитие и формирование мужских половых клеток.
Сперматозоид:
1)мелкая подвижная клетка
2)содержит nc
3)объем ядра больше, чем объем цитоплазмы
4)выделяют головку шейку и хваостик
Ф-ии: передача насл. Инф. от отца к потомкам.
Сперматогенез протекает в извитых канальцах семенников, и его средняя продолжительность от 68 до 75 суток. Сперматогенез у человека начинается с момента полового созревания (Девочки вступают в период полового созревания в 10—12 лет, мальчики в 12—14 лет.) и продолжается в течение всего активного полового периода в больших количествах.
Стадии сперматогенеза:
●размножение;
●рост;
●созревание-деление;
●формирование.
Стадия размножения– стадия интенсивного деления сперматогониев (клетки округлой формы, большое ядро и много цитоплазмы) путем митоза. Большая часть клеток продолжает делиться, а меньшая часть вступает в стадию роста. В этот период клетки растут, накапливают питательные вещества, и на
Стадии роста превращаются в сперматоциты 1-го порядка.
Следующая фаза созревание-деление, характеризуется двумя редукционными делениями, без интерфазы. В результате I-го деления 1 сперматоцит 1-го порядка дает начало 2-м сперматоцитам 2-го порядка, а II-ое деление-созревание приводит к появлению 4 сперматид (овальной формы, меньших размеров)
Фаза формирования происходит в присутствии тестостерона, происходит преобразование сперматид в сперматозоиды. Ядро сперматиды приобретает видоспецифическую форму, хроматин конденсируется. Сперматозоиды– это мелкие, подвижные клетки, размером 30-60 мкм. В сперматозоиде различают головку и хвост.
Головка сперматозоида имеет овоидную форму и включает в себя небольшое плотное ядро, окруженное тонким слоем цитоплазмы. Ядра сперматозоидов характеризуются высоким содержанием нуклеопротаминов и нуклеогистионов. Передняя половина ядра покрыта плоским мешочком, составляющим “чехлик” сперматозоида.
В нем у переднего полюса располагается акросома. Чехлик и акросома являются производными комплекса Гольджи. Акросома содержит набор ферментов, среди которых важное место принадлежит гиалуронидазе и протеазам, способным растворять оболочки, покрывающие яйцеклетку. За головкой имеется кольцевидное сужение.
Головка так же, как и хвостовой отдел, покрыта клеточной мембраной.
Хвостовой отдел сперматозоида состоит из связующих, промежуточных, главной и терминальной частей.
В связующей части или шейке располагаются центриоли – проксимальная и дистальная, от которой начинается осевая нить (аксонема). Промежуточная часть содержит 2 центральных и 9 пар периферических микротрубочек, окруженных расположенными по спирали митохондриями.
Именно митохондрии обеспечивают энергией двигательную активность сперматозоидов, нарушение которой нередко связано с поражением процесса энергообразования в митохондриях. Главная часть по строению напоминает ресничку. Она окружена тонким фибриллярным влагалищем.
Терминальная, или конечная часть содержит единичные сократительные филаменты.
Биологическая роль полового размножения.
При половом размножении наблюдается перекомбинация наследственных признаков родителей, поэтому появляются разнообразные генотипически и фенотипически потомки. Таким образом, половое размножение дает источник изменчивости, благодаря чему появляется возможность лучшего приспособления организмов к среде обитания, к сохранению различных видов организмов.
Источник: https://lektsia.com/3x1b1b.html
Биология и медицина
Развитие яйцеклеток называется оогенезом , а сперматозоидов – сперматогенезом. Процессы развития яйцеклеток и сперматозоидов несколько различаются.
Мужские половые клетки животных – сперматозоиды – образуются в семенниках. В их развитии различают несколько стадий ( рис. 74, слева ). Первая из них – стадия размножения первичных половых клеток – сперматогониев , которые интенсивно делятся путем митоза. Затем эти клетки вступают в профазу I мейоза и превращаются в сперматоциты, первого порядка .
В результате прекращающегося синтеза РНК и, следовательно, белка профазные клетки увеличиваются в размерах. Это – стадия роста. Стадия, во время которой проходят одно за другим два мейотических деления, получила название стадии созревания .
В результате первого деления созревания из одного сперматоцита первого порядка образуется два сперматоцита второго порядка . После второго деления из каждого сперматоцита второго порядка возникают две гаплоидные клетки – сперматиды . Таким образом, из одной исходной клетки, вступившей в мейоз, образуются четыре сперматиды.
Они имеют гаплоидный набор хромосом, но еще не являются специализированными клетками, способными к движению и проникновению внутрь яйцеклетки. Превращение сперматид в сперматозоиды происходит во время следующей стадии – стадии формирования . Она продолжается несколько суток (у человека, например, почти четыре недели).
В это время ядро уменьшается в размерах за счет плотной упаковки хромосом. Резко сокращается объем цитоплазмы, и формируются цитоплазматические структуры, характерные только для сперматозоидов. Рядом с ядром из аппарата Гольджи образуется акросома – пузырек, содержащий ферменты, способные растворить оболочку яйцеклетки в момент оплодотворения.
С противоположной стороны ядра или головки спермия возникает длинный жгут, или хвост, придающий сперматозоиду подвижность. В основании хвоста образующие его фибриллы окружаются слоем митохондрий. Между хвостом и головкой располагается центриоль ( рис. 75 ).
Все эти преобразования обеспечивают дальнейшее поведение сперматозоида: его подвижность и проникновение в яйцеклетку. Плотная упаковка хромосом в головке сперматозоида и отсутствие цитоплазмы позволяют ему хорошо сохранять свою целостность вне организма и делают его очень устойчивым к неблагоприятным воздействиям внешней среды.
Сперматогонии , лежащие непосредственно на базальной мембране извитых семенных канальцев , проходят несколько последовательных стадий митотического деления. Общее количество сперматогоний в яичке человека составляет около 1 млрд.
Различают две основные категории сперматогоний: сперматогонии A и сперматогонии B . Некоторые сперматогонии А, которые делятся митотически, остаются стволовыми , т. е. сохраняют способность к делению и поддерживают свою популяцию.
Остальные дифференцируются в сперматогонии В, которые делятся митотически, дифференцируются в сперматоциты первого порядка и вступают в мейоз .
Сперматоциты первого порядка (первичные), соединены между собой межклеточными мостиками, которые остаются при митозе сперматогоний благодаря неполному разделению клеток. Последующие поколения клеток также остаются соединенными между собой, в результате чего образуется синцитий , клетки которого составляют клон.
Клетки синцития делятся синхронно, лишь единичные клетки не делятся. В результате мейоза I образуются две дочерние клетки сперматоциты второго порядка , которые далее образуют сперматиды.
Таким образом, в результате деления одной сперматогонии образуются четыре сперматиды , каждая из которых обладает гаплоидным набором хромосом рис. 201 .
Дифференцирующиеся сперматиды лежат в углублениях плазматической мембраны клеток Сертоли . При сперматогенезе область ядра, обращенная к стенке семенного канальца, постепенно вытягивается и заостряется.
Комплекс Гольджи формирует гранулы, которые сливаются между собой, образуя акросомальную гранулу (акробласт), контактирующую с апикальной частью ядра.
Одновременно гиперплазирующиеся мешочки комплекса Гольджи окружают акросомальную гранулу, в результате чего образуется акросома , содержащая протеолитические ферменты (в основном гиалуронидазу и трипсиноподобный акрозин ), которые при контакте с яйцеклеткой разрушают ее прозрачную зону.
При отсутствии или недоразвитии акросомы сперматозоид теряет способность оплодотворять яйцеклетку. В процессе сперматогенеза существенно уменьшается количество цитоплазмы формирующегося сперматозоида.
Большая часть ее образует остаточное тельце, которое отделяется от клетки и фагоцитируется сустентоцитом . Меньшая часть цитоплазмы покрывает тонким слоем ядро. Разделение клеток одного клона происходит на последних этапах формирования сперматозоидов, которые отделяются от клеток Сертоли и попадают в просвет канальца, где находится жидкость, вырабатываемая клетками Сертоли .
Сложный процесс сперматогенеза регулируется гормонами.
После полового созревания гипоталамус начинает выделять гонадотропный рилизинг гормон, под влиянием которого гипофиз секретирует фолликулостимулирующий ( ФСГ ) и лютеинизирующий ( ЛГ ) гормоны, воздействующие на клетки Лейдига.
Последние выделяют большое количество тестостерона . Под влиянием клеток фолликулостимулирующего гормона гипофиза клетки Сертоли синтезируют андрогенсвязывающий белок (АСБ) , который переносит мужские половые гормоны к сперматогенным клеткам.
Между процессами образования яйцеклеток и сперматозоидов у млекопитающих существует ряд важных различий. У женщин на ранней стадии эмбриогенеза из размножающихся оогоний образуется ограниченное число ооцитов, которые через определенные промежутки времени по очереди претерпевают овуляцию.
Сперматогенез у мужчин начинается только после полового созревания и затем непрерывно продолжается в эпителиальной выстилке очень длинных, сильно извитых трубочек, называемых семенными канальцами , которые находятся в семенниках.
Незрелые половые клетки – сперматогонии – располагаются на самой периферии канальца, у базальной мембраны, где они все время делятся путем митоза . Некоторые из дочерних клеток перестают делиться и дифференцируются в сперматоциты первого порядка .
Эти клетки вступают, как уже говорилось выше, в I профазу мейоза , в которой происходит кроссинговер между их спаренными гомологичными хромосомами, а затем заканчивают первое деление , образуя по два сперматоцита второго порядка; у человека каждый из них содержит по 22 дуплицированные аутосомы и одну дуплицированную хромосому X или Y.
Каждая хромосома по-прежнему состоит из двух сестринских хроматид , и когда оба сперматоцита второго порядка претерпевают второе деление мейоза , образуются четыре сперматиды с гаплоидным числом одиночных хромосом.
Затем такие гаплоидные сперматиды в результате морфологической дифференцировки превращаются в зрелые спермии, которые выходят в просвет семенного канальца ( рис. 15- 37 и рис. 15-38 ), а позднеее – в придаток семенника, представляющий собой извитую трубочку, охватывающуюю семенник; здесь спермии накапливаются и здесь же продолжается их созревание. ( Browder L., 1980 , Clermont Y., 1972 , Karp G., Berrill N.J., 1981 , Metz C.B., Monroy A., 1985 .)
Процесс образования сперматозоида занимает примерно 70 дней, на 1 г веса семенника образуется примерно 10 млн спермиев в сутки. Эпителий семенного канальца состоит из наружного слоя клеток зачаткового эпителия и примерно шести слоев клеток, образовавшихся в результате многократных делений клеток этого эпителия и соответствуют последовательным стадиям развития сперматозоидов.
Между развивающимися клетками расположены крупные клетки Сертоли, или трофические клетки, расположенные во всем пространстве от наружного слоя канальца до его просвета.
Сперматоциты располагаются в многочисленных впячиваниях на боковых поверхностях клеток Сертоли, здесь они превращаются в сперматиды , а затем переходят на тот край клеток Сертоли, который обращен к просвету семенного канальца , где они созревают, образуя спермии.
Клетки Сертоли обеспечивают созревающим спермиям механическую опору, защиту и питание. Все питательные вещества и кислород , доставляемые развивающимся гаметам по окружающим семенные канальцы кровеносным сосудам и выделяемые в кровь отходы метаболизма проходят через клетки Сертоли. Эти клетки секретируют и жидкость, с которой спермии проходят по канальцам.
Ссылки:
- Интерлокинг
- ПОЛОВАЯ СИСТЕМА МУЖЧИН
- СПЕРМАТОГЕНЕЗ
Источник: http://medbiol.ru/medbiol/reprod/0005dd36.htm
Развитие половых клеток. Сперматогенез
Сперматогенез – процесс развития, созревания и формирования мужских половых клеток. В норме этот период начинается в пубертатном возрасте и продолжается всю жизнь.
Сперматогенез проходит в мужских половых железах – семенниках, это сложная трубчатая железа яйцевидной формы, которая заключена в капсулу – белочную оболочку и состоит из примерно тысячи извитых семенных канальцев, погруженных в соединительную ткань (Рис. 1).
Рис. 1. Семенники
В соединительной ткани содержатся интерстициальные клетки, или клетки Лейдига, в семенных канальцах образуются гаметы сперматозоиды (Рис. 2), а интерстициальные клетки вырабатывают мужской гормон – тестостерон.
Рис. 2. Сперматозоиды и интерстициальные (лейдиговые) клетки
Семенники располагаются вне брюшной полости – мошонке, где температура на 2–3 градуса ниже температуры тела, поэтому сперматозоиды созревают при более низкой температуре, чем температура тела.
Более низкая температура определяется расположением семенников относительно тела и сосудистым сплетением, то есть расположением артерий и вен в семенниках, которые действуют как противоточные теплообменники. Такая температура является обязательным условием для созревания нормальных сперматозоидов.
Сокращение особых мышц перемещает семенники ближе или дальше от тела, чтобы поддерживать в мошонке температуру, оптимальную для развития сперматозоидов.
Если мужчина достиг половой зрелости, но при этом семенники не опустились в мошонку, то развивается состояние, которое носит название крипторхизма, и мужчина навсегда остается стерильным. У мужчин, носящих слишком тесную одежду либо часто принимающих очень горячую ванну, формирование или образование спермиев может очень сильно снизиться и привести к бесплодию.
Фазы сперматогенеза
Сперматогенез включает в себя те же фазы, что и оогенез – фаза размножения, фаза роста и фаза созревания, за которой следует сложный процесс дифференцировки будущих гамет. Образование сперматозоидов занимает 70 суток.
Эпителий семенного канальца (Рис. 3) состоит из наружного слоя клеток зачаткового эпителия и примерно шести слоев клеток, образовавшихся в результате многократных делений клеток этого эпителия, и соответствуют последовательным стадиям развития сперматозоидов.
Рис. 3. Срез семенных канальцев
Клетки зачаткового эпителия делятся и дают начало многочисленным сперматогониям, которые имеют округлую форму и крупное ядро. Сперматогонии содержат диплоидный набор хромосом (46), это фаза размножения.
В фазе роста сперматогонии увеличиваются в размерах и становятся сперматоцитами первого порядка, которые также содержат 46 хромосом.
Эти сперматоциты в результате первого деления мейоза образуют гаплоидные сперматоциты второго порядка, после чего претерпевают второе деление мейоза и превращаются в сперматиды. Сперматоциты и сперматиды содержат гаплоидный набор хромосом – имеют 23 хромосомы. Эта фаза носит название созревания.
В сперматогенезе (Рис.
4) выделяется еще одна фаза – фаза формирования, или спермиогенез, на которой из сперматид формируются сперматозоиды, имеющие определенное строение, хвосты сперматозоидов направлены в просвет канальца, таким образом, из одного диплоидного сперматогония формируется четыре гаплоидных сперматозоида.
Рис. 4. Сперматогенез
Между тяжами из развивающихся клеток располагаются крупные клетки – трофические клетки Сертоли (Рис. 5), которые располагаются во всем пространстве семенного канальца, от наружного слоя до его просвета.
Рис. 5. Клетки Сертоли
Клетки Сертоли выполняют механическую опору, защиту и питание развивающимся сперматозоидам.
Все питательные вещества и кислород, которые доставляются к созревающим гаметам из окружающих семенные канальца кровеносных сосудов, проходят через клетки Сертоли.
Продукты обмена, которые уже не нужны созревающим гаметам, тоже уходят через клетки Сертоли.
Источник: http://100ballov.kz/mod/page/view.php?id=1621
Сперматогенез и овогенез
Сперматогенез и овогенез представляют собой процессы, при которых происходит формирование, рост, а также и созревание мужских и женских половых клеток соответственно. Оба данных явления имеют множество общих сходных черт. Но, несмотря на это, имеются и различия. Давайте подробнее рассмотрим особенности сперматогенеза и овогенеза и охарактеризуем их.
В чем сходства овогенеза и сперматогенеза?
Для начала необходимо отметить, что оба данных процесса имеют одинаковые стадии. Рассмотрим их по порядку:
- Стадия размножения. На данном этапе первичные клетки сперматогонии и оогонии начинают активно делиться путем митоза. При этом необходимо отметить такую особенность данной стадии: у мужчин происходит размножение половых клеток на протяжении всей жизни (с момента половозрелости), а у женщин данная стадия протекает на этапе эмбрионального развития (2-5 месяц развития плода).
- Стадия роста. Происходит сильное увеличение половых клеток в размерах. В итоге они превращаются в сперматоциты и ооциты 1 порядка. При этом ооциты имеют больший размер в виду того, что они накапливают больше питательных веществ, необходимых для развития эмбриона после оплодотворения яйцеклетки.
- Стадия созревания. Характеризуется прохождением мейоз 1 и мейоз 2. В результате первого деления образуются сперматоциты и ооциты 2 порядка, а после второго – зрелые яйцеклетки и сперматиды. Необходимо сказать, что один сперматоцит 1 порядка после деления дает 4 сперматиды, а из ооцита 1 порядка образуется всего одна яйцеклетка и 3 полярных тельца.
В чем различия овогенеза и сперматогенеза?
Проводя сравнительную характеристику овогенеза и сперматогенеза, необходимо сказать, что основным отличием данных процессов является отсутствие в овогенезе 4 стадии формирования. Ее претерпевают исключительно сперматиды, которые в результате превращаются в сперматозоид. Образование данных половых клеток начинается только с наступлением полового созревания у мальчиков.
Все перечисленные выше закономерности сперматогенеза и овогенеза имеют свой биологический смысл. Так, к примеру, неравномерное деление половых клеток при овогенезе способствует образованию лишь одной крупной яйцеклетки с запасом питательных веществ.
Также тот факт, что сперматозоидов образуется намного больше, обусловлен тем, что при оплодотворении яйцеклетки достигает лишь 1 половая клетка мужчины. Остальные погибают еще на пути к женской яйцеклетке.
Предлагаем вам наглядную схему для лучшего понимания процессов сперматогенеза и овогенеза, в которой отображены основные моменты каждого из них.
Источник: https://womanadvice.ru/spermatogenez-i-ovogenez
Сперматогенез – это… что такое сперматогенез?
- сперматогенез — сперматогенез … Орфографический словарь-справочник
- СПЕРМАТОГЕНЕЗ — Процесс образования семенных нитей. Словарь иностранных слов, вошедших в состав русского языка. Чудинов А.Н., 1910. сперматогенез (гр. sperma (spermatos) семя + …генез) процесс образования мужских половых клеток сперматозоидов в семенниках… … Словарь иностранных слов русского языка
- СПЕРМАТОГЕНЕЗ — (от греч. sperma род. п. spermatos семя и …генез), образование дифференцированных мужских половых клеток сперматозоидов; у человека и животных в семенниках, у низших растений в антеридиях. У большинства высших растений в пыльцевой трубке… … Большой Энциклопедический словарь
- СПЕРМАТОГЕНЕЗ — СПЕРМАТОГЕНЕЗ, процесс образования сперматозоидов. СПЕРМАТОЗОИДЫ (спер мин) образуются в несколько стадий деления клеток, которые проходят в семенных канальцах СЕМЕННИКА. Вначале зародышевые клетки превращаются путем деления в сперматогонии,… … Научно-технический энциклопедический словарь
- сперматогенез — сущ., кол во синонимов: 1 • спермиогенез (1) Словарь синонимов ASIS. В.Н. Тришин. 2013 … Словарь синонимов
- СПЕРМАТОГЕНЕЗ — (спермиогенез, sperma topoiesis), процесс развития сперматозоидов или спермий протекает в извитых канальцах яичка, стенки к рых выстланы клеточными элементами, расположенными в несколько слоев (т. н. семенной эпителий) (рис 1) Среди них Рисунок 1 … Большая медицинская энциклопедия
- Сперматогенез — Строение сперматозоида человека Сперматогенез развитие мужских половых клеток (сперматозоидов), происходящее под регулирующим … Википедия
- Сперматогенез — (сперма греч. genesis происхождение, развитие), процесс развития мужских половых клеток, заканчивающийся формированием сперматозоидов. Протекает внутри извитых семенных канальцев, составляющих более 90% объёма яичка взрослого… … Сексологическая энциклопедия
- сперматогенез — [нэ], а; м. [греч. sperma семя и genēs происхождение, возникновение] Процесс образования мужских половых клеток. * * * сперматогенез (от греч. spérma, род. п. spérmatos семя и …генез), образование дифференцированных мужских половых клеток … … Энциклопедический словарь
- сперматогенез — spermatogenesis сперматогенез. Процесс превращения диплоидных первичных половых клеток самцов в гаплоидные дифференцированные половые клетки (сперматозоиды); С. включает 4 основных этапа: размножение (митотическое) сперматогониев, рост… … Молекулярная биология и генетика. Толковый словарь.
Источник: https://dic.academic.ru/dic.nsf/dic_biology/5409/%D0%A1%D0%9F%D0%95%D0%A0%D0%9C%D0%90%D0%A2%D0%9E%D0%93%D0%95%D0%9D%D0%95%D0%97
Презентация биология 10 класс «Развитие половых клеток – Сперматогенез и овогенез»
Слайд 1
Сперматогенез и овогенезПрезентация учителя биологии ГБОУ Школа №879 г. Москвы Титовой С.С.
Слайд 2
Сперматогенез процесс образования мужских половых клеток в половых железах.Этапы сперматогенезаЗона размножения Зона роста Зона созревания Зона формирования
Слайд 3
СперматогенезЗона размножения Диплоидные клетки делятся путем митоза.Происходит увеличение количества клеток.Образуются сперматогонии.Зона роста Происходит рост клеток до размеров, свойственных половым клеткам.Образуются сперматоциты I порядка .2n2n2n2n2n2n2n2n2n2n2n2n2n2n2nМитоз2nСперматогонииСперматоцит I порядка
Слайд 4
СперматогенезЗона созревания Происходит первое деление мейоза.Происходит редукция числа хромосом.Образуются сперматоциты II порядка.Происходит второе деление мейоза. Образуются сперматиды.2nnnnnnnМейозСперматоцит I I порядкаСперматиды
Слайд 5
СперматогенезЗона формирования Сперматиды превращаются в спрематозоиды.Уменьшается ядро.Сокращается объем цитоплазмы.Формируется акросома ( из аппарата Гольджи). Образуется хвост.nnСперматозоиды
Слайд 6
Овогенез процесс образования женских половых клеток в половых железах.Этапы овогенезаЗона размножения Зона роста Зона созревания Зона формирования
Слайд 7
ОвогенезЗона размножения Диплоидные клетки делятся путем митоза.Происходит увеличение количества клеток.Образуются овогонии.Зона роста Происходит рост клеток до размеров, свойственных половым клеткам.Образуются овоцит I порядка .2n2n2n2n2n2n2n2n2n2n2n2n2n2n2nМитоз2nОвогонииОвоцит I порядка
Слайд 8
ОвогенезЗона созревания Происходит первое деление мейоза.Происходит редукция числа хромосом.Образуются овоцит II порядка.Происходит второе деление мейоза. Образуется оотида.2nnnnnМейозовоцит I I порядкаоотида1-е полярное тельце2-е полярное тельце
Слайд 9
Овогенез Зона формирования Ооциты превращаются в яйцеклетки.Полярные тельца исчезают.Происходит синтез митохондрий и рибосом.Развивается гладкая и шероховатая эпс.Синтезируются вещества, которые запасаются в виде желточных и белковых гранул.Сперматозоиды
Слайд 11
Сравнительная характеристика сперматозоидов и яйцеклетокПризнаки для сравнения Сперматозоиды Яйцеклетки Размер клеток Мелкие клетки Крупные клетки Набор хромосом Гаплоидный ( 23 хромосомы) Гаплоидный ( 23 хромосомы) Форма, строение Вытянутая, наличие хвоста Округлая Подвижность Способны к движению Неподвижны Наличие запаса питательных веществ Питательных веществ нет. Запас питательных веществ Функции Доставка генетической информации Развитие зародыша
Источник: http://topslide.ru/biologija/razvitiie-polovykh-klietok–spiermatoghieniez-i-ovoghieniez
Мейоз. Овогенез. Сперматогенез
Содержание
Введение 2
1. Половое размножение многоклеточных 3
2. Гаметогенез 4
2.1 Сперматогенез 4
2.2 Овогенез 5
2.3 Мейоз 6
Вывод 11
Список использованной литературы 12
Введение
Огромное влияние на жизнь человечества в наши дни оказывают генетика и связанные с ней исследования физиологических особенностей размножения человека.
Современные люди часто не помнят или забывают о прошлом и считают, что интерес к генетике и связанным с ней вопросам возник недавно. Но можно утверждать, что интерес к вопросам размножения столь же древен, как и наш собственный биологический вид.
Размножение – основное проявление жизнедеятельности для любого вида, даже если его представители этого и не осознают.
Мендель открыл основные законы наследственности приблизительно в то же время, когда другие биологи принялись изучать строение клетки.
Поэтому когда законы генетики открыли заново в 1900-х годах, ученые уже смогли понять, что так называемые факторы должны соответствовать наблюдаемым в реальности структурам – хромосомам.
Отождествление абстрактных понятий и реальных структур – крупнейшее достижение генетики на ранней стадии ее развития, и оно было сделано в основном благодаря исследованию природы пола. 1
1. Половое размножение многоклеточных
Развитие гамет у многоклеточных животных происходит в половых железах – гонадах (греч. gone – семя). Различают два типа половых клеток: мужские (сперматозоиды) и женские (яйцеклетки). Сперматозоиды развиваются в семенниках, яйцеклетки в яичниках.
Процесс формирования половых клеток (гамет) известен под общим названием гаметогенеза. Он характеризуется рядом весьма важных биологических процессов и протекает несколько по-разному при созревании сперматозоидов (сперматогенез) и яйцеклеток (овогенез). 2
2. Гаметогенез
2.1 Сперматогенез
Семенник состоит из многочисленных канальцев. На поперечном разрезе через каналец видно, что его стенка имеет несколько слоев клеток. Они представляют собой последовательные стадии развития сперматозоидов.
Наружный слой составляют сперматогонии – клетки округлой формы; у них относительно большое ядро и значительное количество цитоплазмы. В период эмбрионального развития и после рождения до полового созревания сперматогонии делятся путем митоза, благодаря чему увеличивается число этих клеток и семенник. Период интенсивного деления сперматогоний называют периодом размножения.
После наступления половой зрелости часть сперматогониев также продолжает делиться митотически и образовывать такие же клетки, но некоторые из сперматогониев перемещаются в следующую зону роста, расположенную ближе к просвету канальца. Здесь происходит значительное возрастание размеров клеток за счет увеличения количества цитоплазмы.
В этой стадии они называются сперматоцидами первого порядка.
Третий период развития мужских гамет называется периодом созревания. В этот период происходят два быстро наступающих одно вслед за другим деления.
Из каждого сперматоцида первого порядка сначала образуются два сперматоцида второго порядка, а затем четыре сперматиды, имеющие овальную форму и значительно меньшие размеры.
Деление клеток во время периода созревания сопровождается перестройкой хромосомного аппарата (происходит мейоз). Сперматиды перемещаются в зону, ближайшую к просвету канальцев, где из них формируются сперматозоиды.
У большинства диких животных сперматогенез происходит лишь в определенные периоды года. В промежутках между ними в канальцах семенников содержатся лишь сперматогонии. Но у человека и большинства домашних животных сперматогенез происходит в течение всего года. 3
2.2 Овогенез
Фазы овогенеза сопоставимы с таковыми при сперматогенезе. В этом процессе также имеется период размножения, когда интенсивно делятся овогонии – мелкие клетки с относительно крупным ядром и небольшим количеством цитоплазмы. У млекопитающих и человека этот период заканчивается еще до рождения.
Сформировавшиеся к этому времени овоциты первого порядка сохраняются далее без изменений многие годы. С наступлением половой зрелости периодически отдельные овоциты вступают в период роста. Овоциты увеличиваются, в них накапливаются желток, жир, пигменты. В цитоплазме клетки в ее органоидах и мембранах происходят сложные морфологические биохимические преобразования.
Каждый овоцит окружается мелкими фолликулярными клетками, обеспечивающими его питание.
Далее наступает период созревания, в процессе которого происходят два последовательных деления, связанных с преобразованием хромосомного аппарата (мейоз).
Кроме того, эти деления сопровождаются неравномерным разделением цитоплазмы между дочерними клетками.
При делении овоцита первого порядка образуется одна крупная клетка – овоцит второго порядка, содержащая почти всю цитоплазму, и маленькая клетка, получившая название полярного, или редукционного тельца.
При втором делении созревания цитоплазма снова распределяется неравномерно. Образуется одна крупная овотида и второе редукционное тельце. В это время первое редукционное тельце также может разделиться на две клетки. Таким образом, из одного овоцита первого порядка образуются одна овотида и три редукционных тельца.
Далее из овотиды формируется яйцо, а редукционные тельца рассасываются или сохраняются на поверхности яйца, но не принимают участия в дальнейшем развитии. Неравномерное распределение цитоплазмы обеспечивает яйцу получение значительного количества цитоплазмы и питательных веществ, которые потребуются в будущем для развития зародыша.
У млекопитающих и человека периоды размножения и роста яйцевых клеток проходят в фолликулах. Фолликул заполнен жидкостью, внутри него находится яйцеклетка. Во время овуляции стенка фолликула лопается, яйцеклетка попадает в брюшную полость, а затем, как правило, в яйцеводы (маточные трубы). Период созревания яйцевых клеток протекает в трубах, здесь же происходит оплодотворение.
У многих животных овогенез и созревание яиц совершаются лишь в определенные сезоны года. У женщин обычно ежемесячно созревает одно яйцо, а за весь период половой зрелости около 400 яиц.
2.3 Мейоз
В ядрах незрелых половых клеток также, как и ядрах соматических клеток, все хромосомы парные, набор хромосом двойной (2 n), диплоидный.
В процессе созревания половых клеток происходит редукционное деление (мейоз), при котором число хромосом уменьшается, становится одинарным (n), гаплоидным. Мейоз (от греч. meiosis – уменьшение) происходит во время гаметогенеза.
Этот процесс совершается во время двух следующих одно за другим делений периода созревания, называемых соответственно первым и вторым мейотическим делением. Каждое из этих делений имеет фазы, аналогичные митозу.
Схематично эти фазы можно изобразить так:
Интерфаза I
Профаза I
Мейоз Деление первое Прометофаза I
Метафаза I
Анафаза I
Телофаза I
Интерфаза II – ин – Профаза II
терокинез Метафаза II
Деление второе Анафаза II
Телофаза II
В интерфазе I (по-видимому, еще в период роста) происходит удвоение количества хромосомного материала путем редупликации молекул ДНК.
Из всех фаз наиболее продолжительна и сложна по протекающим в ней процессам профаза I. В ней различают 5 последовательных стадий. Лептонема – стадия длинных, тонких, слабо спирализованных хромосом, на которых видны утолщения – хромомеры.
Зигонема – стадия попарного соединения гомологичных хромосом, при котором хромомеры одной гомологичной хромосомы точно прикладываются к соответствующим хромомерам другой (это явление называется конъюгацией, или синапсисом). Пахинема – стадия толстых нитей. Гомологичные хромосомы соединены в пары – биваленты.
Число бивалентов соответствует гаплоидному набору хромосом. На этой стадии каждая из хромосом, входящих в бивалент, состоит уже из двух хроматид, поэтому каждый бивалент включает в себя четыре хроматиды.
В это время конъюгирующие хромосомы переплетаются, что приводит к обмену участками хромосом (происходит так называемый перекрест, или кроссинговер). Диплонема – стадия, когда гомологичные хромосомы начинают отталкиваться друг от друга, но в ряде участков, где происходит кроссинговер, они продолжают быть еще связанными.
Диакинез – стадия, на которой отталкивание гомологичных хромосом продолжается, но они еще остаются соединенными в биваленты своими концами, образуя характерные фигуры – кольца и кресты. На этой стадии хромосомы максимально спирализованы, укорочены и утолщены. Непосредственно после диакинеза ядерная оболочка растворяется.
В прометафазе I спирализация хромосом достигает наибольшей степени. Они перемещаются в области экватора.
В метафазе I биваленты располагаются по экватору, так что центромеры гомологичных хромосом обращены к противоположным полюсам и отталкиваются друг от друга.
В анафазе I начинают расходиться к полюсам не хроматиды, а целые гомологичные хромосомы каждой пары, так как в отличие от митоза центромера не делится и хроматиды не разъединяются. Этим первое мейотическое деление принципиально отличается от митоза. Деление заканчивается телофазой I.
Таким образом, во время первого мейотического деления происходит расхождение гомологичных хромосом. В каждой дочерней клетке уже содержится гаплоидное число хромосом, но содержание ДНК еще равно диплоидному их набору. Вслед за короткой интерфазой, во время которой синтеза ДНК не происходит, клетки вступают во второе мейотическое деление.
Профаза II продолжается недолго. Во время метафазы II хромосомы выстраиваются по экватору, центромеры делятся. В анафазе II сестринские хроматиды направляются к противоположным полюсам. Деление заканчивается телофазой II. После этого деления хроматиды, попавшие в ядра дочерних клеток, называются хромосомами.
Итак, при мейозе гомологичные хромосомы соединяются в пары, затем в конце первого мейотического деления расходятся по одной в дочерние клетки.
Во время вторго мейотического деления гомологичные хромосомы расщепляются и расходятся в новые дочерние клетки.
Следовательно, в результате двух последовательных мейотических делений из одной клетки с диплоидным набором хромосом образуются четыре клетки с гаплоидным набором хромосом. В зрелых гаметах количество ДНК вдвое меньше, чем соматических клетках.
При образовании как мужских, так и женских половых клеток, происходят принципиально одни и те же процессы, хотя в деталях они несколько различаются.
Значение мейотического деления в следующем:
Это тот механизм, которым обеспечивается поддержание постоянства числа хромосом. Если бы не происходило редукции числа хромосом при гаметогенезе, то из поколения в поколение возрастало бы их число и был бы утрачен один из существенных признаков каждого вида – постоянство числа хромосом.
При мейозе образуется большое число различных новых комбинаций негомологичных хромосом. Ведь в диплоидном наборе они двойного происхождения: в каждой гомологичной паре одна из хромосом от отца, другая – от матери. Что же происходит при мейозе? Это можно проследить на рис.2.
Изображенные в ядрах сперматогонии и овогонии хромосомы отцовского происхождения обозначены черным цветом, материнские белым. Как видно на рисунке, в сперматозоидах и яйцеклетках они образуют новые сочетания, причем таких сочетаний даже при том же числе хромосом (три пары) получится больше чем изображено.
Следовательно, благодаря такому механизму достигается большое число новых сочетаний наследственной информации, а именно 2, где n – число пар хромосом. Следовательно, у организма, имеющего три пары хромосом, этих сочетаний окажется 2, т.е.8; у дрозофилы, имеющей 4 пары хромосом, их будет 2, т.е.
16, а у человека – 2, что составляет 8388608.
В процессе кроссинговера также происходит рекомбинация генетического материала. Практически все хромосомы, попадающие в гаметы, имеют участки, происходящие как от первоначально отцовских, так и от первоначально материнских хромосом. Этим достигается еще большая степень перекомбинации наследственного материала. В этом одна из причин изменчивости организма, дающий материал для отбора. 4
Вывод
Таким образом, принципиальное отличие полового размножения от бесполого заключается в том, что оно приводит к огромной изменчивости, образованию форм с новыми наследственными свойствами в результате перекомбинации различных свойств обоих родителей. В то время как при бесполом размножении дочерние организмы повторяют наследственную информацию единственной родительской особи. 5
Список использованной литературы
1.Гуттман Б., Гриффитс Э., Сузуки Д., Кулис Т. Генетика. М.: ФАИР – ПРЕСС, 2004., 448 с.
2.Слюсарев А.А. Биология с общей генетикой. М.: Медицина, 1978., 472с.
1
2
Источник: https://doc4web.ru/biologiya/meyoz-ovogenez-spermatogenez.html
СПЕРМАТОГЕНЕЗ
Сперматогенез — это процесс формирования, развития и созревания сперматозоидов из сперматогоний. Он может быть разделен на 2 части: сперматоцитогенез и спермиогенез. Спермацитогенез — процесс формирования сперматоцитов из сперматогоний.
Сперматогонии (Сг) — маленькие клетки, среди которых выделяют тип А и тип В.
- Сперматогонии типа А — клетки с круглым или овальным ядром, содержащим 1 или 2 ядрышка, и со слаборазвитыми органеллами. Ядро и цитоплазма могут быть светлыми (сперматогонии Ас) и темными (сперматогонии Ат). Сперматогонии типа А делятся митозом, при этом одна половина клетки превращается в такую же клетку (чаще типа Ат), а другая постепенно дифференцируется посредством четырех митотических делений в сперматогонии типа В.
- Сперматогонии типа В — грушевидные клетки, узкие основания которых контактируют с базальной мембраной (БМ). Округлое ядро содержит 44 аутосомы и две половые хромосомы (44, XY) и дважды гаплоидный набор ДНК (2п ДНК). Ядрышко обычно находится в центре. Из сперматогонии типа В путем митоза (см. стрелки) появляются две дочерние клетки, которые дифференцируются в первичные сперматоциты (ПС), или сперматоциты I порядка, связанные межклеточными мостиками (ММ).
Сперматоциты I порядка удваивают содержание ДНК (4п ДНК) и вступают в первое (редукционное) мейотическое деление, профаза I которого состоит из следующих 4 стадий:
- лептотена: материнская и отцовская (т. е. гомологичные) хромосомы сближаются. На рисунке они показаны серым и черным цветами. Каждая хромосома состоит из 2 хроматид (Хр), обозначенных номерами 1, 2, 3 и 4 соответственно. Гомологичные хромосомы фиксированы к ядерной оболочке (ЯО) прикрепляющими площадками (ПП). Каждая хромосома имеет центромеру (Ц) и белковую ось (Ос). Половые хромосомы (X и Y) также удваивают содержание своей ДНК;
- зиготена: характерно тесное сближение гомологичных хромосом, называемое синапсом. Этот процесс начинается с момента, когда концы хромосом прикрепляются вместе к ядерной оболочке. Синаптонемальный комплекс (СК) начинает формироваться между концами хромосом и затем стремительно продолжается в направлении, указанном стрелкой, по типу застежки «молния», выравнивая две гомологичные хромосомы таким образом, что их гены располагаются друг против друга. Каждая хромосома состоит из двух тесно прилежащих друг к другу хроматид, поэтому она называется бивалентом, а структура в целом из двух совмещенных хромосом — тетрадой;
- пахитена: в так называемых рекомбинанионных узелках (РУ), расположенных на определенных интервалах вдоль синаптонемального комплекса, происходит кроссинговер между двумя несестринскими хроматидами, при этом они обмениваются между собой целыми сегментами;
- диплотена: синаптонемальный комплекс расходится и гомологичные хромосомы с центромерами (Ц) отдаляются друг от друга на некоторое расстояние, в то время как биваленты остаются объединенными одной или несколькими хиазматами (Хи), представляющими собой места, где еще продолжается перекрест.
- Диакинез и мейотическая метафаза I: хромосомы конденсируются в более толстые и короткие и отходят от ядерной оболочки, которая постепенно исчезает. Каждая тетрада, состоящая из 4 хроматид (Хр), хорошо видна в световом микроскопе. Сестринские хроматиды объединяются своими центромерами, которые теперь развиваются в кинетохоры (К), в то время как несестринские хроматиды соединены своими хиазматами. Хорошо видны измененные сегменты несестринских хроматид. Микротрубочки веретена деления (БД) прикрепляются к кинетохорам сестринских хроматид.
- Анафаза I: после деполимеризации трубочек веретена деления биваленты отделяются друг от друга и перемещаются к противоположным полюсам клетки. В результате этого деления образуется два вторичных сперматоцита, или сперматоцита II порядка. Кроссинговер между хромосомами X и Y также имеет место, но для ясности процесса он не изображен.
В результате первого (редукционного) мейотического деления появляется два вторичных сперматоцита (ВС), или сперматоцита II порядка, также называемых пресперматидами, каждый из которых содержит гаплоидный набор хромосом (22 X; 22 Y) и половину ДНК первичного сперматоцита (2п ДНК).
Сперматоциты II порядка вскоре вступают во второе (эквационное) мейотическое деление, но без S-фазы клеточного цикла, т. е. без репликации ДНК в интерфазе.
Во время метафазы II кинетохоры (К) делятся и мигрируют к противоположным сторонам сестринских хроматид (X).
Укорочение микротрубочек веретена деления (ВД) отделяет сестринские хроматиды, теперь рассматриваемые как хромосомы, друг от друга в течение анафазы и, в результате формируя 4 сперматиды (Ст), соединенные межклеточными мостиками (ММ).
Эти клетки также содержат гаплоидный набор хромосом (22Х; 22Y), но только половину ДНК от вторичных сперматоцитов (п ДНК). В ходе спермиогенеза сперматиды превращаются без дополнительного деления в сперматозоиды (С). Только на последней стадии их развития межклеточные мостики разрушаются и сперматозоиды отделяются друг от друга.
Таким образом, второе (эквационное) мейотическое деление похоже на нормальный митоз с разницей в том, что в клетке имеется только половиннный набор хромосом.
К сожалению, невозможно увидеть отдельные хромосомы в ядре сперматозоида, поскольку они сильно конденсированы. Только в дидактических целях на рисунке изображены серые хромосомные сегменты в ядре сперматозоида, чтобы показать генетическую разницу между различными сперматозоидами.
Профаза I продолжается 16—24 дня. Около 64—74 дней необходимо для развития сперматогонии в зрелые сперматиды. Поскольку перенос сперматозоидов через придаток яичка занимает 8—17 дней, полная продолжительность сперматогенеза до эякуляции зрелых сперматозоидов составляет примерно 82 дня.
Сперматогенез контролируется гипоталамогипофизарным комплексом: гонадотропин-рилизинг-гормон стимулирует гонадотропные клетки гипофиза высвобождать фолликулостимулирующий (ФСГ) и лютейнизирующий (ЛГ) гормоны. Они активизируют не только клетки Сертоли, т. е.
развитие сперматогенных клеток, но также рост семенных канальцев и через адениловую циклазо-циклическую АМФ систему — секрецию андроген-связываюшего белка клетками Сертоли.
Лютеинизирующий гормон стимулирует клетки Лейдига, продуцирующие тестостерон, необходимый для созревания сперматозоидов.
Тестостерон ингибирует секрецию гонадотропин-рилизинг-гормона с помощью механизма отрицательной обратной связи, так же, как и фолликулостимулирующего и лютеинизирующего гормонов. Ингибин — вещество, секретируемое клетками Сертоли, избирательно снижает секрецию фолликулосгимулирующего гормона и, возможно, также гонадотропин-рилизинг-гормона.
На рисунке изображены 2 типа сперматозоидов:
– сперматозоид с Х-хромосомой, так называемый гиносперматозоид.
– сперматозоид, несущий Y-хромосому, — андросперматозоид.
Х-хромосома больше Y-хромосомы, поэтому гиносперматозоид имеет примерно на 7 % большую поверхность головки. В результате андросперматозоид несколько подвижнее и имеет более высокую возможность оплодотворения вторичного ооцита.
Это может отчасти объяснить, почему на каждые 100 девочек рождаются 106 мальчиков.
Источник: http://tardokanatomy.ru/content/spermatogenez
Сперматогенез
Направление дифференциации мужских половых клеток резко отличается от стратегии формирования женских гамет.
Дифференциация мужских гамет предполагает максимальную редукцию цитоплазматического тела и развитие двигательного аппарата. Объем спермия определяется практически размерами компактизованного ядра, нескольких митохондрий, центросомы и величиной жгутика.
Следует обратить внимание на то, что развитие мужских и женских гамет, столь различающееся по дифференциации клеточных структур, происходит в соответствии с одной и той же принципиальной схемой ядерных преобразований, в ходе которых диплоидные первичные половые клетки (ППК) превращаются в гаплоидные гаметы.
Первые описания подвижных живчиков («зверьков», лат. animalculi) в семени животных принадлежат И. Гаму и А. Левенгуку (1677). Аналогичные образования были обнаружены и в семени человека, сперматозоид которого был увековечен на известном рисунке Николаса Хартсекера (1694).
Стоявший на позициях анималькулизма Хартсекер предполагал, что именно в этих подвижных клетках заложены в миниатюре все структуры взрослого организма, отвергая тем самым гипотезы о паразитарной природе живчиков. Термин сперматозоид был введен в научную литературу К. Бэром (1817).
Окончательное решение вопроса о природе сперматозоидов как мужских половых клеток принадлежит Альберту Кёлликеру (1817-1905), который на нескольких десятках видов беспозвоночных доказал развитие спермиев из клеток семенников.
Кёлликер не только продемонстрировал все переходные формы от недифференцированной ППК до высокоспециализированного спермия, но и установил видовую специфичность спермиев и даже обнаружил у некоторых видов полиморфизм спермиев.
Тем не менее явление редукции числа хромосом в ходе сперматогенеза оставалось незамеченным вплоть до последней четверти XIX столетия, когда Ван Бенеден, исследуя развитие мужских и женских половых клеток аскариды, описал мейоз (van Beneden, 1884). Новая страница в изучении структуры и функции спермиев была открыта в 50-е годы XX столетия, когда с использованием электронной микроскопии обнаружились принципиально новые особенности тонкой организации половых клеток.
По структурной организации и способу функционирования сперматозоиды можно подразделить на несколько типов. У подавляющего большинства видов сперматозоиды жгутиковые. Такие сперматозоиды обычно имеют три четко выраженных отдела: головку, в которой расположено ядро, среднюю часть, или шейку и жгутик.
Известны некоторые группы животных, у которых спермии не имеют жгутиков. Безжгутиковые сперматозоиды характерны для некоторых Ecdysozoa. Так, у круглых червей сперматозоиды амебоидные.
Своеобразные безжгутиковые спермии обнаружены и у многих членистоногих, их спермии округлой или звездчатой формы могут перемещаться с помощью псевдоподий или остаются неподвижными.
Сперматозоиды, как правило, микроскопических размеров. Спермин гидры имеют жгут длиной до 30 мкм. У некоторых книдарий его длина достигает 90 мкм. Известны и гигантские спермин. Например, у Drosophila bifurca длина сперматозоида составляет 58 мм. В этом случае спермий в 20 раз длиннее тела взрослой особи и в 15 000 раз длиннее спермия человека.
Хроматин ядра спермия сильно конденсирован и плотно упакован. В процессе формирования сперматозоида объем его ядра резко уменьшается (например, у петуха от 110 до 2 мкм3). Конденсация хроматина обусловлена замещением гистонов, характерных для соматических клеток, переходными белками, а затем и особыми высокоосновными спермийными белками — протаминами и иными аргининбогатыми белками.
Жгутиковые сперматозоиды обычно имеют акросому (от греч. акрос — крайний, сора — тело), хотя у многих животных, таких, как гидра, некоторые моллюски, костистые рыбы, акросома отсутствует.
Акросома представляет собой секреторный пузырек, который формируется при участии аппарата Гольджи и содержит разнообразные гидролитические ферменты. У многих животных, в частности у млекопитающих, акросомный пузырек в процессе формирования спермия принимает вид шапочки, облегающей переднюю часть ядра.
У иглокожих между акросомным пузырьком и ядром имеется субакросомное пространство, в котором локализуются молекулы глобулярного актина.
Митохондрии часто сливаются между собой, образуя крупную хондросферу. Они обычно локализуются в основании жгутика вокруг центриоли и, как полагают, генерируют энергию для движения жгутика.
Иногда, например в сперматидах насекомых, моллюсков и кольчатых червей, слившиеся митохондрии располагаются в виде «побочного ядра» сбоку от ядра.
Тем не менее, в конечном счете, митохондриальное побочное ядро окружает аксонему жгутика, или располагается рядом с ним.
Центриоль, которая в период делений сперматогониев и сперматоцитов определяет формирование веретена, в ходе спермиогенеза преобразуется в базальное тельце жгутика.
Эта трансформация центриоли дрозофилы происходит при участии центросомина, особого белка центросомы.
Мутация mfs, при которой синтез центросомина не происходит, вызывает нарушение спермиогенеза и, как следствие этого, стерильность самцов.
Основу жгутика составляет так называемый осевой комплекс, или аксонема, состоящая из двух центральных одиночных микротрубочек, вокруг которых расположены девять дублетов микротрубочек.
Кроме этого комплекса спермин многих видов имеют еще одно внешнее кольцо, также состоящее из девяти фибрилл и придающее жгутику необходимую жесткость.
Иногда в заднем отделе спермия образуется ундулирующая мембрана.
До настоящего времени не разгадано явление полиморфизма спермиев: у ряда животных в семеннике развиваются спермин двух разновидностей.
Например, у некоторых гастропод (Prosobranchia) наряду с типичными жгутиковыми, или эупиренными сперматозоидами, обладающими оплодотворяющей способностью, развиваются короткие червеобразные апиренные формы, часто с аномальным гаплоидным набором хромосом. В других случаях апиренные спермин имеют два жгутика. Доля апиренных сперматозоидов постоянна для данного вида.
У ряда американских сумчатых наблюдается феномен попарного соединения зрелых спермиев, поступивших в эпидидимис. При этом лежащая над акросомой плазматическая мембрана одного сперматозоида плотно соединяется с соответствующей областью другого. Предполагается, что это объединение спермиев, жгутики которых расположены асимметрично относительно ядра, улучшает их подвижность.
Число спермиев, выводимых при осеменении, огромно. Так, у опоссума оно достигает 13 млн, у кролика их на порядок больше. У человека ежедневно может формироваться более 100 млн спермиев.
Естественно, что этот интенсивный процесс требует координации пролиферации стволовых клеток и включения программ сперматогенеза, контролирующих преобразование диплоидных сперматогониев в зрелые гаплоидные сперматозоиды.
Сперматогенез подразделяется на несколько фаз. Первая из них — фаза размножения, в течение которой образующиеся после деления стволовых клеток сперматогонии митотически делятся. Число митотических циклов у разных видов варьирует от 1 до 14. Вторая фаза — фаза коммитации, в течение которой включаются генетические программы инициации сперматогенеза. Третья фаза — фаза созревания.
Этот этап охватывает премейотический синтез ДНК и два мейотических деления. Мужские половые клетки, находящиеся в первом мейотическом цикле, называют первичными сперматоцитами (сперматоциты первого порядка), а во втором — вторичными сперматоцитами (сперматоциты второго порядка).
После второго мейотического деления образуется гаплоидная сперматида и наступает заключительная фаза — фаза формирования спермия. В ходе спермиогенеза происходит резкое уменьшение объема ядра в результате элиминации нуклеоплазмы и конденсации хроматина. Изменение структуры хроматина в первую очередь обусловлено заменой соматических гистонов специфическими спермийными.
В этот период образуется жгутик и завершается формирование акросомы. Характерной особенностью сперматогенеза является неполный цитокинез в период митотических делений сперматогоний и первого мейотического деления сперматоцитов. В результате потомки одного сперматогония образуют синцитиальную цисту.
Полагают, что наличие цитоплазматических мостиков, связывающих сперматоциты, обеспечивает высокую синхронность дифференциации мужских половых клеток, входящих в состав одной цисты.
Не исключено, что синцитиальная структура важна для поддержания фенотипической эквивалентности цитоплазмы сперматид, что особенно важно в отношении иРНК половых хромосом, так как многие белки, важные для спермиогенеза, контролируются генами X и Y хромосом. Так, например, описан перенос иРНК через межклеточные мостики гаплоидных сперматид мыши с помощью белка TB-RBP (testis-brain-RNA-binding protein).
Индивидуализация мужских гамет осуществляется на завершающем этапе спермиогенеза, когда происходит «сбрасывание» цитоплазмы вместе со всеми органеллами — рибосомами, эндоплазматической сетью, аппаратом Гольджи. У некоторых животных эта остаточная, или резидуальная (от лат. residuum — остаток) цитоплазма, отделяющаяся от головки спермия, фагоцитируется клетками фолликулярного эпителия семенника.
В опытах по так называемому микрооплодотворению, когда мужские половые клетки вводятся в ооцит с помощью микроинъекции или путем искусственного «сшивания» гамет, выяснилось, что уже ранние сперматиды и даже вторичные и иногда первичные сперматоциты обладают потенцией к образованию зиготы, способной к дальнейшему развитию. Другими словами, сложные процессы цитодифференциации, происходящие в период спермиогенеза, необходимы прежде всего для образования аппарата движения, формирования механизмов заякоривания и слияния с женской гаметой.
Процесс сперматогенеза предполагает тесную корреляцию митотических и мейотических циклов со сложной программой клеточной дифференциации.
О существовании относительной автономии программы мейоза свидетельствует мутация twine, вызывающая у дрозофилы существенные аномалии мейоза и вместе с тем не затрагивающая заметным образом дифференциацию сперма, тид.
Несмотря на нарушения сегрегации мейотических хромосом и цитокинеза, в семенниках мутантных самцов наблюдаются половые клетки, у которых происходит конденсация хроматина, характерное видоизменение формы ядра и образование жгутика.
Наряду с этим известны мутации, которые блокируют одновременно и мейоз и спермиогенез. Эти мутации свидетельствуют о том, что на определенной стадии сперматогенеза действует сопряженная программа развития, в равной степени необходимая как для мейотического цикла, так и для дифференциации сперматид.
Семенник позвоночных подразделен на два отдела: семенные канальцы и межканальцевое пространство, или соединительнотканную строму. В последней находится обширная сеть кровеносных сосудов, интерстициальные клетки или клетки Лейдига, а также макрофаги, фибробласты, лимфоциты и тучные клетки.
Семенные канальцы отделены от стромы так называемой ограничивающей мембраной, которая образована уплощенными клетками, расположенными в 2-3 слоя. Полагают, что эти клетки могут превращаться в клетки Лейдига.
Эпителий семенных канальцев имеет сложный состав и образован клетками двух типов: половыми (гоноциты) и соматическими клетками фолликулярного эпителия. Последние часто называют клетками Сертоли, или опорными клетками. Реже их называют сустентоциталш (от лат. sustentas — поддерживающий).
Между половыми и опорными клетками в течение всего сперматогенеза (у крысы цикл сперматогенеза длится 42 дня, у человека 74 дня) сохраняется постоянная взаимосвязь. Клетки Сертоли многофункциональны.
Они обеспечивают питание дифференцирующихся мужских половых клеток и способствуют созданию необходимой для нормального сперматогенеза концентрации андрогенов. Опорные клетки играют также важную роль в перемещении половых клеток из базального отсека канальца в околополостной.
Они участвуют в создании гематотестикулярного барьера, который предохраняет половые клетки от разного рода метаболитов и токсинов, а также предотвращает автоиммунизацию организма против собственных спермиев. Благодаря строго скоординированной сборке и разборке плотных контактов между клетками Сертоли происходит своего рода «шлюзование» сперматоцитов в околополостной компартмент канальца с сохранением непроницаемой границы между отсеками.
Функциональная активность семенника у позвоночных во многом определяется гормональной регуляцией со стороны гипоталамо-гипофизарной системы. Инициация сперматогенеза обусловлена секрецией лютеинизирующего гормона (ЛГ) и фолликулстимулирующего гормона (ФСГ), которые вырабатываются в передней доле гипофиза.
В канальцах мишенями ФСГ являются клетки Сертоли, а в строме — макрофаги. Действие ЛГ на сперматогенез опосредовано клетками Лейдига: под действием ЛГ в этих клетках активируется синтез мужского полового гормона тестостерона, наличие которого является непременным условием сперматогенеза.
В обоих случаях происходит выработка паракринных (т. е. действующих вблизи от места секреции) факторов, поддерживающих активность клеток Лейдига. ФСГ стимулирует также аккумуляцию тестостерона в клетках Сертоли, создавая предпосылку нормального сперматогенеза.
Дифференциация клеток Сертоли у мыши по времени совпадает с экспрессией гена Tsx, что дает основание предполагать участие этого гена в осуществлении специфической функции клеток Сертоли — обеспечении сперматогенного цикла у животных, достигших половой зрелости.
Для поддержания этой функции необходимы и белки, продуцируемые клетками Лейдига. Показано, что эти белки (Gas 6, S) служат лигандами, которые воспринимаются специфическими тирозинкиназными рецепторами клеток Сертоли.
Поступающие извне сигналы инициируют активность генетических программ сперматогенеза. Методом иммуноблотинга было показано, что у мыши в период сперматогенеза в результате экспрессии генов в половых клетках появляется целый ряд специфических для этого периода антигенов.
Глобальная транскрипция прекращается за несколько дней до завершения спермиогенеза. В период спермиогенеза активируется трансляция ранее синтезированных иРНК. В частности, активируется трансляция транскриптов протамина.
Благодаря динамической модификации хроматина и точной координации процессов транскрипции и процессинга иРНК достигается экспрессия белков, специфических для разных этапов спермиогенеза.
Решающую роль в активации генов, ответственных за сперм иогенез и экспрессирующихся в гаплоидном геноме, играет цАМФ-зависимый транскрипционный фактор CREM. Оказалось, что необходимым элементом для экспрессии генов в период спермиогенеза является активация аденилатциклазной системы мужских половых клеток.
Было показано, что гонадотропные гормоны гипофиза, взаимодействуя с рецепторами мембран, активируют G-белок и связанную с ним аденилатциклазу. Это в конце концов индуцирует накопление цАМФ в клетках Сертоли, воспринимающих сигнал ФСГ, и в клетках Лейдига, взаимодействующих с ЛГ.
Повышение уровня цАМФ вызывает диссоциацию неактивного комплекса протеинкиназы А на две субъединицы — регуляторную и каталитическую. После диссоциации каталитическая субъединица фосфорилирует ядерные транскрипционные факторы класса bZIP, которые связываются со специфической нуклеотидной последовательностью TGACGTCA в промоторах генов-мишеней. Этот сайт называют CRE (от сАМР responsive element).
Соответственно транскрипционные факторы, связывающиеся с CRE, называются CREB (CRE binding protein) и CREM (CRE modulator). Оказалось, что эти факторы играют важную роль в спермиогенезе. У мышей половое созревание сопровождается появлением больших количеств фактора CREM. Ген crem экспрессируется в сперматоцитах начиная со стадии пахитены.
Синтез же соответствующего белка происходит только на стадии ранней сперматиды. У сперматозоидов белок CREM отсутствует. Наблюдения на нокаутированных мышах crem показали, что такие мыши не имеют видимых отклонений от нормы за исключением изменения семенников, масса которых на 20-25% меньше массы семенников нормальных мышей.
Гомозиготные самцы crem в отличие от самок были стерильными: в семенной жидкости сперматозоиды отсутствовали. При микроскопическом исследовании семенных канальцев оказалось, что сперматогенез у мышей, лишенных гена crem, не шел дальше стадии ранней сперматиды. Спермиогенез был полностью подавлен, хотя клетки Сертоли имели нормальный вид.
У дрозофилы во время спермиогенеза экспрессируется специфический для мужских половых клеток ген don juan.
Начало синтеза белка Don Juan приходится на стадию удлиненной сперматиды и продолжается в течение всего спермиогенеза. Этот белок молекулярной массой 29 кДа характеризуется высоким содержанием лизина.
Как полагают, белок Don Juan участвует в дифференциации митохондрий и обеспечивает морфогенез жгутика спермия.
Важный момент в дифференциации сперматид — экспрессия специфических для сперматозоидов трансмембранных белков, которые обеспечивают адгезию мужской гаметы и ооцита.
У млекопитающих эту функцию выполняют β-1,4-галактозил-трансфераза, зонадгезин и фертилин.
Последний принадлежит семейству белков ADAM (a disintegrin and metalloprotease), одна из функций которых состоит в процессинге рецепторов плазматической мембраны.
Источник: http://www.activestudy.info/spermatogenez/