Спорогенез и гаметогенез у растений
Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап — спорогенез — завершается образованием гаплоидных клеток — спор, в ходе 2-го этапа — гаметогенеза — происходит ряд делений гаплоидных клеток, прежде чем образуются зрелые гаметы.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) — мега- или макроспорогенезом. Микроспорогенез протекает аналогично делению созревания у животных мужских половых клеток до стадии сперматиды, а мегаспорогенез — соответственно до стадии незрелой яйцеклетки — ооцита II.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У животных после двух мейотических делений формируются гаметы, и никаких дополнительных клеточных делений не происходит.
У растений в результате двух мейотических делений возникает гаплоидная спора, из которой развивается гаметофит, представляющий собой у низших растений (грибов, печеночников, мхов, ряда водорослей) целый организм и наиболее продолжительную стадию цикла существования.
У высших растений гаплоидная фаза редуцирована, однако ядра мужской и женской спор претерпевают ряд митотических делений, прежде чем образуются гаметы.
Микроспорогенез и микрогаметогенез
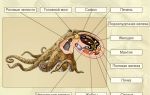
Мы рассмотрим микроспорогенез и микрогаметогенез на примере покрытосеменных растений как наиболее общем. В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делении становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
Схема микроспорогенеза (1-6) и микрогаметогенеза (5-10) у растений
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
У однодольных растений каждое деление ядра в мейозе, как правило, сопровождается цитокинезом; у двудольных оба деления клетки наступают одновременно по окончании мейоза.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, кутинизированная, поверхность ее либо гладкая, либо шероховатая; приспособленная для переноса пыльцы и прилипания ее к рыльцу пестика.
Этим заканчивается микроспорогенез вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся.
В ней накапливаются запасные питательные вещества, который в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые в отличие от сперматозоидов животных называются спермиоклетками, или спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: Два из них — спермии и одно — вегетативное. При образовании пыльцевой трубки это вегетативное ядро в полужидком диффузном состоянии переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены С. Г. Навашиным в 1910 г. на лилейных растениях.
Мегаспорогенез и мегагаметогенез
У покрытосеменных растений женский гаметофит — это зародышевый мешок, который закладывается и развивается внутри семяпочки.
Схема мегаспорогенеза (1-5) и мегагаметогенеза (6-12) у растений
Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез. В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоры.
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной.
Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития), судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем ее ядро претерпевает ряд эквационных делений. При этом сама клетка не делится, делится только ядро.
У разных систематических групп растений число эквационных делений ядра мегаспоры может варьировать от одного до трех.
У большинства растений (70% видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других — в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка, и две так называемые синергиды образуют яйцевой аппарат. Однако из этих трех клеток после оплодотворения развивается только одна, а две другие разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца.
Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное или центральное, ядро зародышевого мешка. Это ядро с цитоплазмой зародышевого мешка называют обычно центральной клеткой зародышевого мешка. Однако часто полярные ядра, передвинувшиеся к центру, не сливаются до оплодотворения.
Оставшиеся у халазального конца зародышевого мешка три ядра также обособляются в клетки; они называются антиподами.
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 наследственно одинаковых гаплоидных ядер, из которых только одно дает яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно.
В одних случаях, как мы только что рассмотрели, развитие зародышевого мешка начинается из одной гаплоидной споры (моноспорический тип развития), в других — из двух (биспорический тип) и четырех спор (тетраспорический тип).
Типы развития зародышевых мешков
Как мы указывали, при моноспорическом типе развивается лишь одна мегаспора из четырех, а остальные три разрушаются подобно тому, что имеет место с редукционными тельцами у животных. При других типах развития зародышевого мешка сохраняется разное количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям.
Образование пыльцевых зерен и зародышевых мешков у цветковых растений
Изучая гаметогенез, нельзя не поражаться тому параллелизму, который наблюдается при созревании половых клеток у животных и растений, несмотря на то, что их расхождение (дивергенция) в филогенезе произошло на очень раннем этапе возникновения клеточной организации. Это указывает на однотипность принципов построения ряда приспособительных механизмов как в растительном, так и животном мире.
Итак, изучение развития половых клеток у животных и у растений показало, что формирование гамет является сложным процессом. Прежде чем яйцеклетка и спермий объединятся в процессе оплодотворения, они претерпевают ряд превращений.
Однако половые клетки так же, как и клетки любой другой ткани, происходят из соматических. Поэтому их нельзя рассматривать как нечто обособленное от тела организма. Вместе с тем половые клетки имеют и свои особенности.
Основными характерными моментами, отличающими их от соматических клеток, являются следующие:
1. У разных животных и растений на разных стадиях дифференциации тканей зародыша происходит обособление половых клеток. Процесс закладки и дифференциации, половых клеток у животных называется зачатковым путем.
2. В процессе развития половых клеток особое значение имеет мейоз с характерными для него стадиями деления ядра, а именно профазой I, во время которой конъюгируют гомологичные хромосомы, метафазой I и анафазой I, когда осуществляется редукция числа хромосом и расхождение гомологичных хромосом к различным полюсам.
3. Главным свойством половых клеток является способность их при оплодотворении сливаться в одну с образованием зиготы, которая претерпевает затем дробление и развитие. Соматические клетки этой способностью, как правило, не обладают.
Источник: http://www.activestudy.info/sporogenez-i-gametogenez-u-rastenij/
Спорогенез и гаметогенез у растений
Содержание
1. Спорогенез и гаметогенез у растений
2. Норма реакции генотипа
3. Бесплодие отдаленных гибридов, его причины и способы преодоления
Список литературы
1. Спорогенез и гаметогенез у растений
Способность к размножению, или самовоспроизведению, является одним из обязательных и важнейших свойств живых организмов.
Размножение поддерживает длительное существование вида, обеспечивает преемственность между родителями и их потомством в ряду многих поколений. Оно приводит к увеличению численности особей вида и способствует его расселению.
У растений, подавляющее большинство которых ведет прикрепленный образ жизни, расселение в процессе размножения – единственный способ занять большую территорию обитания.
Различают два типа размножения: бесполое и половое. В, бесполом размножении участвует только одна родительская особь, которая делится, почкуется и образует споры. Размножение при помощи вегетативных органов у растений называется вегетативным. В случае полового размножения особи нового поколения появляются при участии двух организмов – материнского и отцовского.
Вегетативное размножение основано на способности организмов восстанавливать (регенерировать) недостающие части. Этот способ размножения широко распространен в природе, но с наибольшим разнообразием оно осуществляется у растений, особенно у цветковых.
Бесполое размножение характеризуется тем, что для воспроизводства потомства образуются специализированные клетки – споры, каждая из которых прорастает и дает начало новому организму.
Спорообразование встречается у простейших (малярийный плазмодий), грибов, водорослей, мхов, плаунов, хвощей и папоротников.
У голо – и покрытосеменных растений споры непосредственно в процессе, размножения не участвуют.
Споры образуются путем митоза или мейоза в обычных вегетативных клетках материнского организма или специальных органах – спорангиях и представляют собой микроскопические одноклеточные образования.
При любой форме бесполого размножения – частями тела или спорками – наблюдается увеличение численности особей данного вида без повышения их генетического разнообразия: все особи являются точной копией материнского организма.
Эта особенность используется человеком для получения однородного, с хорошими признаками, потомства у плодово-ягодных, декоративных и других групп растений. Новые признаки у таких организмов появляются только в результате мутаций.
Половое размножение существенно отличается от бесполого тем, что в данном случае генотип потомков возникает в результате перекомбинации генов, принадлежащих обоим родителям. Это повышает возможности организмов в приспособлении к меняющимся условиям среды.
Половое размножение характеризуется наличием полового процесса, одним из важнейших этапов которого является слияние половых клеток, или гамет, специализированных гаплоидных клеток, одетых плазматической мембраной.
Гаметы различаются по строению и физиологическим свойствам и делятся на мужские (подвижные – сперматозоиды, неподвижные – спермин) и женские (яйцеклетки). В отличие от спор одна гамета, за исключением случаев партеногенеза, не может дать начало новой особи.
Этому предшествует процесс слияния двух половых клеток – оплодотворение, в результате которого образуется зигота. В дальнейшем из зиготы развивается зародыш нового организма.
Образование половых клеток (гаметогенез) у водорослей, многих грибов и высших споровых растений происходит путем митоза или мейоза в специальных органах полового размножения: яйцеклеток – в оогониях или архегониях, сперматозоидов и спермиев – в антеридиях. В процессе формирования половых клеток выделяют три стадии – размножения, роста и созревания.
Первичные половые клетки делятся путем митоза (период размножения), в результате чего их количество постоянно возрастает. В период роста деление клеток прекращается, и они начинают усиленно расти.
При этом будущие яйцеклетки (ооциты) увеличиваются в размерах иногда в сотни и даже в тысячи раз за счет накопления в их цитоплазме запасных питательных веществ в виде желтка. Размеры незрелых мужских гамет (сперматоцитов) увеличиваются незначительно.
Затем происходит их мейотическое деление, что приводит к образованию четырех гаплоидных клеток. При сперматогенезе все четыре клетки в дальнейшем превращаются в сперматозоиды.
Половое размножение растений включает несколько физиологических процессов: цветение, опыление, оплодотворение и образование плода и семени.
При оплодотворении пыльца, попадая на рыльце пестика, прорастает, образуя пыльцевую трубку, достигающую семяпочки в завязи цветка. У растений может быть одна семяпочка или несколько. В нижней части пыльцевой трубки образуются спермин.
Затем происходит так называемое двойное оплодотворение, при котором один из спермиев сливается с яйцеклеткой, а другой – с центральной клеткой семяпочки.
После оплодотворения из яйцеклетки в результате многократного деления развивается зародыш семени, а из оплодотворенной центральной клетки образуется запас питательных веществ семени.
Так в результате двойного оплодотворения образуется семя, состоящее из оболочки, зародыша и запаса питательных веществ, из которого впоследствии развивается новое растение. Полученное при половом размножении потомство наследует признаки обоих родителей.
2. Норма реакции генотипа
Источник: https://www.studsell.com/view/168106/
Читать контрольная по биологии: “Спорогенез и гаметогенез у растений”
(Назад)(Cкачать работу)
Функция “чтения” служит для ознакомления с работой. Разметка, таблицы и картинки документа могут отображаться неверно или не в полном объёме!
Содержание 1. Спорогенез и гаметогенез у растений 2 Список литературы 15
1. Спорогенез и гаметогенез у растений
Способность к размножению, или самовоспроизведению, является одним из обязательных и важнейших свойств живых организмов.
Размножение поддерживает длительное существование вида, обеспечивает преемственность между родителями и их потомством в ряду многих поколений. Оно приводит к увеличению численности особей вида и способствует его расселению.
У растений, подавляющее большинство которых ведет прикрепленный образ жизни, расселение в процессе размножения – единственный способ занять большую территорию обитания.
Различают два типа размножения: бесполое и половое. В, бесполом размножении участвует только одна родительская особь, которая делится, почкуется и образует споры. Размножение при помощи вегетативных органов у растений называется вегетативным. В случае полового размножения особи нового поколения появляются при участии двух организмов – материнского и отцовского.
Вегетативное размножение основано на способности организмов восстанавливать (регенерировать) недостающие части. Этот способ размножения широко распространен в природе, но с наибольшим разнообразием оно осуществляется у растений, особенно у цветковых.
Бесполое размножение характеризуется тем, что для воспроизводства потомства образуются специализированные клетки – споры, каждая из которых прорастает и дает начало новому организму.
Спорообразование встречается у простейших (малярийный плазмодий), грибов, водорослей, мхов, плаунов, хвощей и папоротников.
У голо – и покрытосеменных растений споры непосредственно в процессе, размножения не участвуют.
Споры образуются путем митоза или мейоза в обычных вегетативных клетках материнского организма или специальных органах – спорангиях и представляют собой микроскопические одноклеточные образования.
При любой форме бесполого размножения – частями тела или спорками – наблюдается увеличение численности особей данного вида без повышения их генетического разнообразия: все особи являются точной копией материнского организма.
Эта особенность используется человеком для получения однородного, с хорошими признаками, потомства у плодово-ягодных, декоративных и других групп растений. Новые признаки у таких организмов появляются только в результате мутаций.
Половое размножение существенно отличается от бесполого тем, что в данном случае генотип потомков возникает в результате перекомбинации генов, принадлежащих обоим родителям. Это повышает возможности организмов в приспособлении к меняющимся условиям среды.
Половое размножение характеризуется наличием полового процесса, одним из важнейших этапов которого является слияние половых клеток, или гамет, специализированных гаплоидных клеток, одетых плазматической мембраной.
Гаметы различаются по строению и физиологическим свойствам и делятся на мужские (подвижные – сперматозоиды, неподвижные – спермин) и женские (яйцеклетки). В отличие от спор одна гамета, за исключением случаев партеногенеза, не может дать начало новой особи.
Этому предшествует процесс слияния двух половых клеток – оплодотворение, в результате которого образуется зигота. В дальнейшем из зиготы развивается зародыш нового организма.
Образование половых клеток (гаметогенез) у водорослей, многих грибов и высших споровых растений происходит путем митоза или мейоза в специальных органах полового размножения: яйцеклеток – в оогониях или архегониях, сперматозоидов и спермиев – в антеридиях. В процессе формирования половых клеток выделяют три стадии – размножения, роста и созревания1.
Первичные половые клетки делятся путем митоза (период размножения), в результате чего их количество постоянно возрастает. В период роста деление клеток прекращается, и они начинают усиленно расти.
При этом будущие яйцеклетки (ооциты) увеличиваются в размерах иногда в сотни и даже в тысячи раз за счет накопления в их цитоплазме запасных питательных веществ в виде желтка. Размеры незрелых мужских гамет (сперматоцитов) увеличиваются незначительно.
Затем происходит их мейотическое деление, что приводит к образованию четырех гаплоидных клеток. При сперматогенезе все четыре клетки в дальнейшем превращаются в сперматозоиды.
Половое размножение растений включает несколько физиологических процессов: цветение, опыление, оплодотворение и образование плода и семени.
При оплодотворении пыльца, попадая на рыльце пестика, прорастает, образуя пыльцевую трубку, достигающую семяпочки в завязи цветка. У растений может быть одна семяпочка или несколько. В нижней части пыльцевой трубки образуются спермин.
Затем происходит так называемое двойное оплодотворение, при котором один из спермиев сливается с яйцеклеткой, а другой – с центральной клеткой семяпочки. После оплодотворения из яйцеклетки в результате многократного деления развивается зародыш семени, а из оплодотворенной центральной клетки образуется запас питательных веществ семени.
Так в результате двойного оплодотворения образуется семя, состоящее из оболочки, зародыша и запаса питательных веществ, из которого впоследствии развивается новое растение. Полученное при половом размножении потомство наследует признаки обоих родителей2. 2.
Норма реакции генотипа При формировании генетических представлений о связи между геном и признаком изначально предполагалось, что каждому признаку соответствовал особый детерминант (наследственный фактор), который обусловливал развитие своего признака.
Однако такие представления далеки от истины, а прямые и однозначные связи гена с признаком на самом деле скорее исключение, чем правило. Было установлено, что на один признак могут влиять многие гены и, наоборот, один ген часто влияет на многие признаки. Кроме того, действие гена может быть изменено соседством других генов или условиями внешней среды.
В онтогенезе действуют скорее не отдельные гены, а весь генотип как целостная интегрированная система со сложными связями и взаимодействиями ее компонентов. Более того, эта система не является застывшей, она динамична, меняется, совершенствуется во времени, в результате генных мутаций постоянно появляются новые гены.
Могут формироваться также качественно новые хромосомы за счет хромосомных мутаций и даже новые геномы за счет геномных мутаций. Вновь возникшие гены могут сразу же вступать во взаимодействие с уже имевшимися генами или менять, модифицировать характер работы последних, даже будучи рецессивными, т.е.
не проявляясь сами по себе3.
Таким образом, в каждый конкретный промежуток времени у каждого вида растений и животных генотип проявляет себя как исторически сложившаяся к данному моменту целостная система.
Характер проявления действия гена может изменяться в различных ситуациях и под влиянием различных факторов.
Законы Менделя отражают законы наследования, то есть передачи генов в ряду поколений, только при обязательном соблюдении двух условий: гены должны быть локализованы в разных парах гомологичных хромосом (это дает им возможность независимо комбинироваться и наследоваться) и за каждый признак должен отвечать только один ген. Однако это далеко не всегда так. Для того, чтобы убедиться в том, что характер проявления генов разнообразен, рассмотрим свойства генов и особенности их проявления в признаках:
ген дискретен в своем действии, то есть, прерывист, обособлен в своей активности от других генов;
ген специфичен в своем проявлении, т.е. отвечает за строго определенный признак (на молекулярном уровне каждый ген отвечает за синтез одного конкретного белка);
ген может действовать градуально, то есть может усиливать степень проявления признака (например, увеличивать количество синтезируемого вещества) при увеличении числа доминантных аллелей (дозы гена);
один ген может влиять на развитие разных признаков – это множественное, или плейотропное, действие гена;
разные гены могут оказывать одинаковое действие на развитие одного и того же признака – это множественные гены, или полигены; при этом чаще всего наблюдается усиление или ослабление признаков – в таком случае это кумулятивное (накопительное) действие гена, которое
Источник: https://referat.co/ref/179011/read?p=1
Спорогенез и гаметогенез у растений.Двойн. оплодотв. У покрытосеменных
Поиск Лекций
Процесс формирования полов. клеток у растен. подраздел. на 2 этапа:1) Спорогенез-завершаеться образованием гаплоидных клеток-спор,») гаметогенез-происходит ряд делений гаплоидных клеток,прежде чем образуються зрелые гаметы.Процесс образов. микроспор назыв-микроспорогенезом,а процесс образ мегаспор-макроспорогенезом.
Микроспорогенез протекает аналогично сперматогенезу у животных, вплоть до образования сперматид, а мегаспорогенез соответствует образованию ооцита И.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У растений отсутствует зачатковый путь в том смысле, как это имеет место у животных.
В субэпидермальной ткани молодого пыльника, называемой археспорием, каждая клетка после ряда делений становится материнской клеткой пыльцы, которая проходит все фазы мейоза.
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Они лежат четверками, их называют тетрадами спор.
При созревании тетрады распадаются на отдельные микроспоры. Этим заканчивается микроспорогенез.
Вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, не способны к движению и называются спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три клетки. Две из них — спермии и одна вегетативная.
Мегаспорогенез и мегагаметогенез. В субзпидермальном слое молодой семяпочки обособляется археспориальная клетка, часто только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоу.
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады гаплоидна. Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития). Судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти, и затем ее ядро претерпевает ряд митозов. При этом сама клетка не делится, она образует зародышевый мешок.
У разных растений число митозов может варьировать от одного до трех. У большинства растений (70% видов покрытосеменных) бывает три деления, в результате возникает восемь одинаковых ядер.
Во время этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других в противоположном конце зародышевого мешка, называемого халазальным. В дальнейшем ядра обособляются в самостоятельные клетки, имеющие значительное количество цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка и две так называемые синергиды — образуют яйцевой аппарат. Синергиды играют вспомогательную роль при оплодотворении, они скоро разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца.
Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное, или центральное, ядро зародышевого мешка. Оставшиеся у халазального конца зародышевого мешка три ядра обособляются в клетки; они называются антиподами. Антиподы, так же как и синергиды, играют вспомогательную роль при развитии зиготы и вскоре разрушаются.
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 одинаковых гаплоидных ядер, из которых только одно образует яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно.
Кроме моноспорического типа развития, который только что рассмотрен, существуют и другие. При различных типах развития зародышевого мешка сохраняется разное'1 количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям (если сохраняются две мегаспоры — биспорический; четыре — тетраспорический тип развития).
В завязь покрытосеменных растений проникает два спермия, один из них сливается с яйцеклеткой, дав начало диплоидному зародышу. Другой соединяется с центральной диплоидной клеткой. Образуется триплоидная клетка, из которой возникнет эндосперм – питательный материал для развивающегося зародыша .
Этот процесс, характерный для всех покрытосеменных, открыт в конце прошлого века С.Г. Навашиным и получил название двойного оплодотворения. Значение двойного оплодотворения,заключается в том, что обеспечивается активное развитие питательной ткани уже после оплодотворения.
Поэтому семяпочка у покрытосеменных не запасает питательных веществ впрок и, следовательно, развивается гораздо быстрее, чем у многих других растений, например у голосеменных.
15.Цитологический механизм формирования гамет у моногетерозиготы.Вероятностный характер сочетания гамет при оплодотворении.Решетка Пеннета. Если данный организм, будь то животное или растение, имеет два совершенно одинаковых гена, например два гена черной окраски (BB) или два гена коричневой (bb), то его называют гомозиготным, или «чистым», по данному признаку.
Если же он содержит один доминантный и один рецессивный ген (например, Bb), то его называют гетерозиготным, или гибридным. Используя эти термины, мы можем дать более полные определения доминантному и рецессивному генам: рецессивный ген — это ген, проявляющийся только в гомозиготном состоянии, доминантный же ген оказывает свое действие как в гомозиготном, так и в гетерозиготном состоянии.
Когда у черных морских свинок F1 (Bb) образуются гаметы, хромосома с геном B и гомологичная ей хромосома с геном b конъюгируют, а затем расходятся, так что каждый сперматозоид и каждая яйцеклетка получают либо ген B, либо ген но никогда не могут получить оба гена одновременно.
Понятно, что у гетерозиготных особей Bb сперматозоиды и яйцеклетки двух типов — с геном B и с геном b — образуются в равном числе. Но если имеется два типа яйцеклеток и два типа сперматозоидов, то при оплодотворении возможны четыре комбинации гамет.
Между яйцеклетками и сперматозоидами, содержащими одинаковые гены, нет ни какого-то особого притяжения, ни отталкивания, так что все эти четыре комбинации равновероятны. Вероятности соответствующих комбинаций можно определить путем алгебраического умножения.
(1/2 B + 1/2 b) X (1/2 B + 1/2 b) = (1/4 BB + 1/4 Bb + 1/4 bB + 1/4 bb)=3/4 с домин и 1/4рецес.
яйцеклетки сперматозоиды сочетание при оплодотворении
Можно также наглядно представить все возможные комбинации, построив так называемую «решетку Пённета», вдоль верхней стороны которой выписывают все типы яйцеклеток, а вдоль левой стороны — все типы сперматозоидов, клетки же заполняют всеми возможными их сочетаниями в зиготе.
Три четверти всех потомков будут иметь генотип BB или Bb и потому будут иметь черную окраску, а у одной четверти будет генотип bb, и эти особи будут коричневыми.
Многочисленные эксперименты показали, что генетический механизм, первоначально постулированный Менделем для объяснения результатов гибридизации Горохов, в равной мере действует при скрещивании самых различных животных и растений.
Источник: https://poisk-ru.ru/s22607t2.html