Концепция данной теории заключается в том, что передача наследственной информации в ряду поколений осуществляется путем передачи хромосом, в которых в определенной линейной последовательности расположены гены.
Данная теория была сформулирована в начале XX века. Значительный вклад в ее развитие внес американский генетик Томас Морган.

Рекомендую осознать и запомнить следующие положения хромосомной теории:
- Гены расположены в хромосомах в линейном порядке
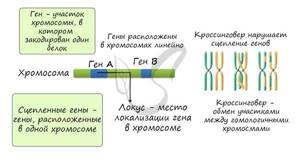
- Каждый ген занимает в хромосоме определенное место – локус
- Гены, расположенные в одной хромосоме, образуют группу сцепления
- Сцепление генов может нарушаться в результате кроссинговера
- Частота кроссинговера между генами прямо пропорциональна расстоянию между ними
- Расстояние между генами измеряется в морганидах (1 морганида – 1% кроссинговера)
Группы сцепления
В предыдущей статье были раскрыты суть и применение в задачах III закона Менделя, закона независимого наследования, в основе которого лежат гены, расположенные в разных хромосомах. Но что если гены лежат в одной хромосоме? Такие гены образуют группу сцепления, в этом случае говорят о сцепленном наследовании.
Группа сцепления – совокупность всех генов, расположенных в одной хромосоме, вследствие чего они наследуются совместно. Число групп сцепления равно гаплоидному набору хромосом: у женщины 23 группы сцепления (23 пара – половые хромосомы XX), а у мужчины – 24 группы сцепления (X и Y представляют собой две отдельные группы).
Сцепление генов
Томас Морган в своих экспериментах изучал наследование признаков плодовых мушек дрозофил: серый (A) – черный (a) цвет тела, длинные (B) – зачаточные (b) крылья. В первом эксперименте Морган скрестил чистые линии плодовых мушек: серых с длинными крыльями (AABB) и черных с зачаточными (aabb).
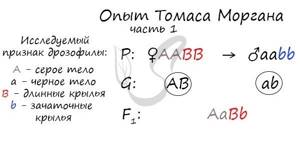
Только что вы видели первый закон Менделя (единообразия) в действии, правда, в несколько ином варианте – при дигибридном скрещивании. Но суть та же: в первом поколении все особи получаются единообразны по исследуемому признаку, с генотипом AaBb – с серым телом и длинными крыльями.
Далее Морган применил анализирующее скрещивание. Полученную в первом поколении дигетерозиготу (AaBb) он скрестил с черной особью с зачаточными крыльями (aabb). Результат весьма удивил Моргана и его коллег: помимо потомства с ожидаемыми фенотипами (серое тело + длинные крылья, черное тело + зачаточные крылья) были получены особи со смешанными признаками.

Потомство со смешанными признаками подразумевает под собой особи Aabb (серое тело + зачаточные крылья) и aaBb (черные тело + длинные крылья). Но откуда они могли взяться, если гены A и B находятся в одной хромосоме? Значит, образовались еще какие-то дополнительные гаметы, помимо AB и ab?
Объясняя полученные в потомстве фенотипы, которые содержали смешанные признаки, Томас Морган пришел к выводу, что между гомологичными хромосомами произошел кроссинговер, в результате которого образовались гаметы Ab, aB – кроссоверные гаметы.
Очевидно, что в данном случае расстояние между генами A и B было 17 морганид, так как каждой кроссоверной гаметы (соответственно и особей) образовалось по 8.5%. Не забывайте, что процент кроссинговера равен расстоянию между генами. Поскольку расстояние было 17 морганид = 17%, то на каждую из кроссоверных гамет приходится половина – 8.5%
Пример решения генетической задачи №1
“Катаракта и полидактилия у человека обусловлены доминантными аутосомными генами, расположенными в одной хромосоме. Гены полностью сцеплены. Какова вероятность родить здорового ребенка в семье, где муж нормален, жена гетерозиготна по обоим признакам, мать жены также страдала обеими аномалиями, а отец был нормален”.
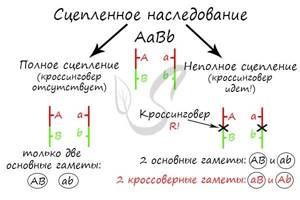
Очень важно обратить внимание на то, что “гены полностью сцеплены” – это говорит об отсутствии кроссинговера, и то, что мы заметили это, обеспечивает верное решение задачи.

Самое главное, что вам следует усвоить: поскольку гены полностью сцеплены (кроссинговер отсутствует), женщина с генотипом AaBb может образовать только два типа гамет – AB, ab. Кроссоверные гаметы (Ab, aB) не образуются. Всего возможных генотипов потомков получается два, из которых здоров только один – aabb. Шанс родить здорового ребенка в такой семье ½ (50%).
Пример решения генетической задачи №2
“Гены доминантных признаков катаракты и эллиптоцитоза локализованы в 1-й аутосоме. Гены неполностью сцеплены. Женщина, болеющая катарактой и эллиптоцитозом, отец которой был здоров, выходит замуж за здорового мужчину. Определите возможные фенотипы потомства и вероятность рождения больного обеими аномалиями ребенка в этой семье”.
Ключевые слова в тексте этой задачи, на которые следует обратить внимание: “гены неполностью сцеплены”. Это означает, что между ними происходит кроссинговер.
Генотип женщины остается неясен из текста задачи. Раз она больна, то он может быть: AaBb, AABB, AABb, AaBB. Однако в тексте дано то, что развеет сомнения: “отец которой был здоров”. Если ее отец был здоров, то его генотип был aabb, значит он передал дочери гамету ab. Теперь становится очевидно, что генотип дочери AaBb – она дигетерозиготна.

В данном случае между генами A и B произошел кроссинговер, их сцепление нарушилось. В результате образовались кроссоверные гаметы Ab, aB – которые привели к образованию особей с со смешанными признаками (Aabb, aaBb). Вероятность рождения в этой семье ребенка, больного обеими аномалиями, составляет ¼ (25%).
Наследование, сцепленное с полом
Половые хромосомы X и Y определяют пол человека. Генотип XX характерен для женщин, а XY – для мужчин. Мужская Y-хромосома не содержит аллелей многих генов, которые есть в X-хромосоме, вследствие этого наследственными заболеваниями, сцепленными с полом, чаще болеют мужчины.
Природа, несомненно, бережет женских особей.
Женщины имеют две гомологичные хромосомы XX, и если ген наследственного заболевания попал в одну из X-хромосом, то чаще всего в другой X-хромосоме окажется “здоровый” ген, доминантный, которой подавит действие рецессивного гена. С генетической точки зрения, женщина будет носительницей заболевания, может его передать по поколению, но сама болеть не будет.
У мужчин если ген заболевания оказался в X-хромосоме, то не проявиться он не может. Именно по этой причине мужчины чаще страдают дальтонизмом, гемофилией и т.д.
Не у всех организмов особь мужского пола характеризуется набором хромосом XY, а женского – XX. У пресмыкающихся, птиц, бабочек женские особи имеют гетерогаметный пол- XY, а мужские – XX. То же самое относится к домашним курам: петух – XX, курица – XY.
Решим несколько задач по теме наследования, сцепленного с полом. Речь в них будет идти о сцепленных с полом признаками – признаками, гены которых лежат не в аутосомах, а в гетеросомах (половых хромосомах).
Пример решения генетической задачи №3
“Рецессивный ген дальтонизма располагается в X-хромосоме. Женщина с нормальным зрением (отец был дальтоник) выходит замуж за мужчину с нормальным зрением, отец которого был дальтоником. Определите возможные фенотипы потомства”.
Подробности о родословной важны и помогают заполнить белые пятна. Если отец женщины был дальтоником (XdY), то очевидно, что он передал ей хромосому Xd, так как от отца дочери всегда передается X-хромосома.
Значит женщина гетерозиготна по данному признаку, а у мужчины возможен лишь один вариант здорового генотипа – XDY. То, что его отец был дальтоником несущественно, ведь отец всегда передает сыну Y-хромосому.
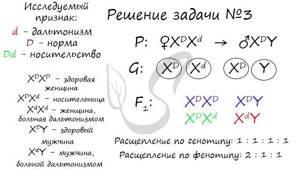
Возможные фенотипы потомства:
- XDXD, XDXd – фенотипически здоровые девочки
- XDY – здоровый мальчик
- XdY – мальчик, который болен дальтонизмом
Пример решения генетической задачи №4
“Гипоплазия зубной эмали наследуется как сцепленный с X-хромосомой доминантный признак, шестипалость – как аутосомно-доминантный. В семье, где мать шестипалая, а у отца гипоплазия, родился пятипалый здоровый мальчик. Напишите генотипы всех членов семьи по данным признакам. Возможно ли у них рождение ребенка с двумя аномалиями одновременно?”
Ответ на вопрос: “Каковы генотипы матери и отца?” – лежат в потомстве. Пятипалый здоровый мальчик имеет генотип aaXbY. Чтобы сформировался такой генотип, от матери должна прийти гамета aXb, а от отца – aY. Выходит, что единственно возможный генотип матери – AaXbXb, а генотип отца – aaXBY.
Рождение ребенка с двумя аномалиями возможно – AaXBXb, вероятность такого события ¼ (25%).
Пример решения генетической задачи №5
“Рецессивные гены, кодирующие признаки дальтонизма и гемофилии, сцеплены с X-хромосомой. Мужчина с нормальным цветовым зрением и гемофилией женится на здоровой женщине, отец которой был дальтоником, но не гемофиликом. Известно, что мать женщины была гомозиготна по исследуемым признакам. Какое потомство получится от брака их дочери со здоровым мужчиной?”
Генотип мужчины вопросов не вызывает, так как единственный возможный вариант – XhDY. Генотип женщины дает возможность узнать ее отец (XHdY), который передал ей гамету XHd (отец всегда передает дочке X хромосому, а сыну – Y), следовательно, ее генотип – XHDXHd
Как оказалось, возможны два варианта генотипа дочери: XHDXhD, XHdXhD. Генотип здорового мужчины XHDY. Следуя логике задачи, мы рассмотрим два возможных варианта брака.
Не забывайте, что на экзамене схема задачи не является ответом. Ответ начинается только после того, как вы напишите слово “Ответ: …”. В ответе должны быть указаны все фенотипы потомства, их описание, что возможно покажется рутинными при большом числе потомков, но весьма приятным, если вы верно решили задачу и получили за нее заслуженные баллы 🙂
Сцепленное наследование признаков. Хромосомная теория наследственности
После того как Грегор Мендель открыл единые закономерности
наследственности.
В начале 20-го века
учёные генетики стали проводить множество экспериментов по скрещиванию на самых
различных объектах. В итоге обнаружилось, что закономерности, установленные Менделем проявляются не всегда.
Мендель скрещивал дигетерозиготы – организмы которые отличались по двум
признакам. Признаки, которые рассматривал Мендель были локализованы в разных
гомологичных хромосомах.
- Вспомним что третий закон
Менделя формулируется так: каждая пара аллельных генов (и альтернативных
признаков, контролируемых ими) наследуется независимо друг от друга. - Что значит независимо
друг он друга? - При скрещивании организмов
1-го поколения при мейозе образуются 4 типа гамет.
Где гены комбинируются в
различных сочетаниях. Такие сочетания получились потому что гены находились в
разных хромосомах.
Но в 1906 году Уильям Бэтсон и Риджинальд Пэннет, проводя скрещивание растений душистого горошка и
анализируя наследование нескольких признаков формы пыльцы и окраски цветков,
обнаружили, что эти признаки не дают независимого распределения в потомстве в
соотношении 3:1, гибриды всегда повторяли признаки родительских форм.
Стало ясно, что не для
всех признаков характерно независимое распределение в потомстве и свободное
комбинирование.
Дело в том, что ген формы
пыльцы и ген окраски цветка лежат в одной хромосоме.
Признаков в организме
значительно больше чем хромосом, в которых эти признаки локализованы.
Следовательно, каждая хромосома несёт не один ген, а целую группу генов,
отвечающих за развитие разных признаков.
Изучением наследования
признаков, гены которых локализованы в одной хромосоме, занимался Томас Морган.
Он предложил закон
сцепленного наследования (закон Моргана): гены, которые находиться
в одной хромосоме, при мейозе попадают в одну гамету, то есть наследуются
сцеплено.
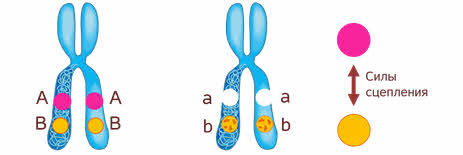
Что значит сцеплено? То
есть между генами, которые находиться в одной хромосоме возникают силы
сцепления, то есть силы взаимодействия. И чем ближе эти гены, тем сильнее
взаимодействие.
- Если Мендель проводил
свои опыты на горохе, то для Моргана основным объектом стала фруктовая мушка
дрозофила, которая имела диплоидный набор из 8 хромосом. - Небольшие размеры,
короткий жизненный цикл и простота культивирования позволяет использовать ряд
видов дрозофил как образцовые объекты генетических исследований. - Самец и самка внешне
хорошо различимы — у самца брюшко меньше и темнее.
Дрозофила фруктовая —
наиболее важный для научных исследований вид дрозофил. Главными её
характеристиками как модельного объекта является малое число хромосом.
Дрозофила каждые две недели при температуре 25 °С достаточно легко размножаются
в пробирках и даёт многочисленное потомство.
Рассмотрим один из первых
экспериментов Томаса Моргана по изучению сцепленного наследования.
Скрещивая мушку дрозофилу
с серым телом и нормальными крыльями с мушкой, имеющей темную
окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды,
которые имели серое тело и нормальные крылья.
- Ген А-большое отвечает за
серое тело, рецессивный ген а-малое за чёрную окраску тела, доминантный ген В-большое за развитие длинных крыльев, а рецессивный ген
бэ-малое за не развитие крыльев, то есть крылья остаются в зачаточном
состоянии. - Значит ген, определяющий
серую окраску брюшка, доминирует над темной окраской, а ген, обуславливающий
развитие нормальных крыльев, — над геном недоразвитых. - Далее при скрещивании
двух гетерозигот которые имеют серое тело и длинные
крылья в первом поколении получается два фенотипических класса организмов.
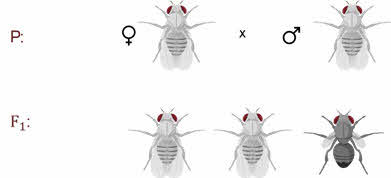
Которые имеют серое тело
и длинные крылья и чёрное тело с зачаточными крыльями.
То есть здесь расщепление
идёт именно на 2 класса, а не на четыре как при дигибридном
скрещивании Менделя.
Почему два? Дело
в том, что гены окраски тела и длинны крыльев сцеплены в хромосоме.
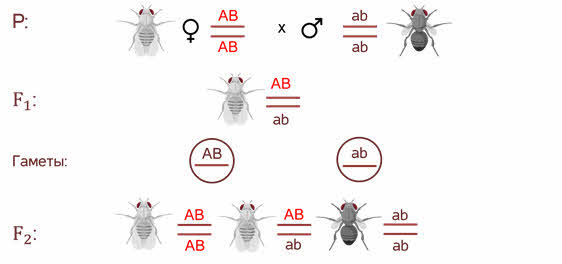
Символы АB АB ab аb располагаются не рядом как мы
записывали их ранее, а как бы друг под другом с двумя чёрточками. Чёрточками мы
условно обозначаем хромосомы.
В первом поколении
организм гетерозиготен по обоим генам, но при
образовании гамет эта гетерозиготность не даёт всех
возможных комбинаций. То есть родительские гены остаются связанные между собой
и гаметы получаются 2х типов.
- При комбинации такого
рода гамет в потомстве возникают всего 3 генотипических класса потомков. - Морган исследуя
наследование сцепленных генов обнаружил что существует нарушение этого правила
по дигибридному скрещиванию Менделя. - Он провёл анализирующее
скрещивание.
Взял дигетерозиготную
особь, которая получилась при скрещивании в первом поколении и скрестил её с
чёрной мушкой с зачаточными крыльями, то есть оба рецессивных признака. У него
получился необычный результат.
Морган рассуждал.
Если
гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном
скрещивании должны были получиться две группы особей, повторяющие признаки родительских
форм, так как материнский организм должен образовывать гаметы только двух типов
— АB
и аb, а отцовский — один тип — аb.
Следовательно, в потомстве должны образовываться две группы особей, имеющих
генотип АB аb
и
аa BB.
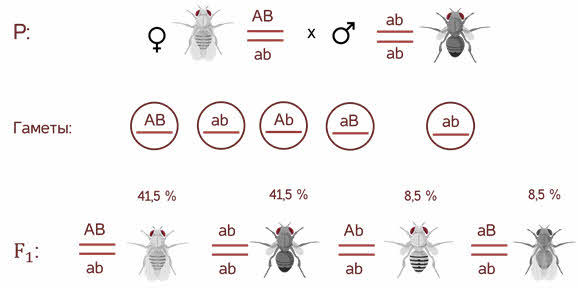
- Однако в потомстве
появляются особи (пусть и в незначительном количестве) с перекомбинированными
признаками, то есть имеющие генотип Аa bb и aa Bb. - В потомстве явно
преобладали особи с признаками родительских форм (41,5% были серые длиннокрылые
и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек
имела иное, чем у родителей, сочетание признаков (8,5% были серые с зачаточными
крыльями и 8,5% — черные длиннокрылые). - Такие результаты могли
быть получены только в том случае, если гены, отвечающие за окраску тела и
форму крыльев, находятся в одной хромосоме. - В профазе первого мейотического деления гомологичные хромосомы (то есть
идентичные хромосомы одной пары) конъюгируют (сближаются), и могут разрываться
в месте контакта в этот момент между ними может произойти обмен участками –
кроссинговер.
Кроссинго́вер
или перекрёст — это процесс обмена участками гомологичных хромосом во время
конъюгации в профазе первого мейоза. В результате образуются крассоверные гаметы.
Организмы, которые
возникают в результате слияния кроссоверных гамет
называются рекомбинантными.
Так в результате,
кроссинговера в некоторых клетках происходит обмен участками хромосом между
генами А и В, появляются гаметы Аb и аB, и, как следствие, в потомстве образуются четыре группы
фенотипов, как при свободном комбинировании генов.
Однако кроссинговер
происходит не после каждой конъюгации (сближения хромосом). И определить
в каких участках хромосом он произойдет достаточно
сложно.
В ходе эксперимента
Томасу Моргану удалось доказать, что частота кроссинговера между генами
прямо пропорциональна расстоянию между ними в хромосоме. То есть можно
сказать что, чем дальше гены находятся друг от друга в хромосоме, тем чаще
между ними происходит кроссинговер.
Если рассматривать 2 гена
А и В мы можем увидеть 2 случая.
В первом случае гены А и В находиться по разные стороны перекрёста. Тогда после
прохождения кроссинговера мы увидим новые сочетания аллелей этих двух генов. В
данном случае Аb
и аB.
Во 2м случае гены А и B находиться
по одну сторону от перекрёста. Тогда после прохождения кроссинговера новых
сочетаний аллелей данных двух генов мы не увидим.
- Таким образом существуют
понятия полного и неполного сцепления. - Неполное сцепление — это
разновидность сцепленного наследования, при котором гены анализируемых
признаков располагаются на некотором расстоянии друг от друга, что делает
возможным кроссинговер между ними. - Полное сцепление — это
разновидность сцепленного наследования, при котором гены анализируемых
признаков располагаются так близко друг к другу, что кроссинговер между ними
становится невозможным.
Это открытие позволило
лаборатории Томаса Моргана разработать метод. Который позволяет построить хромосомные
карты.
- Хромосомные карты
— это графическое изображение хромосомы, на котором определенные
локусы (гены) отмечены соответственно расстоянию между ними. - Хромосомные карты
составляют при помощи генетического анализа, который позволяет точно определить
местоположение в хромосоме любого гена. - Хромосомная теория
наследственности - Изучение Морганом
наследования родительских признаков гибридами дрозофилы показало, что число
групп сцепленного наследования было равным количеству пар гомологичных хромосом.
Например, у человека 46
хромосом, следовательно, 23 группы сцепления. У дрозофилы 8 хромосом, то есть 4
группы сцепления.
На этом основании был
сделан вывод о строгой локализации конкретных генов в определенных
парах хромосом.
Возникновение кроссоверных (рекомбинантных) особей дрозофилы можно было
объяснить только линейным расположением генов в хромосомах и их обменом при
кроссинговере в профазе первого мейоза.
Томас Морган обосновал
хромосомную теорию наследственности. Согласно этой теории, передача
наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены.
Таким образом, именно хромосомы представляют собой материальную основу
наследственности.
Формированию хромосомной
теории способствовали данные, полученные при изучении генетики пола, когда были
установлены различия в наборе хромосом у организмов различных полов.
Хромосомная теория
наследственности сформулирована в 1911 г. американским учёным Томасом Морганом.
Её сущность заключается в следующем:
- ·
Основным материальным носителем
наследственности являются хромосомы с локализованными в них генами. - ·
Гены в хромосомах расположены линейно,
каждый ген имеет определенное место (локус) в
хромосоме; - ·
Гены,
расположенные в одной хромосоме, образуют группу сцепления и наследуются
совместно; - ·
Число групп сцепления равно гаплоидному
набору хромосом у гомогаметных особей и n+1 у гетерогаметных
особей. - ·
Между гомологичными хромосомами может
происходить обмен участками (кроссинговер); в результате кроссинговера
возникают гаметы, хромосомы которых содержат новые комбинации генов. - ·
Сцепление генов может нарушаться в
результате кроссинговера;
·
Частота кроссинговера между гомологичными
хромосомами зависит от расстояния между генами, локализованными в одной
хромосоме. Чем это расстояние больше, тем выше частота кроссинговера.
Значение этой теории
заключается в том, что она дала объяснение законам Менделя, вскрыла
цитологические основы наследования признаков и генетические основы теории
естественного отбора.
Лекция № 18. Сцепленное наследование
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков.
Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган.
Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.
1 — некроссоверные гаметы; 2 — кроссоверные гаметы.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — и , а отцовский — один тип — .
Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип и . Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип и . Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз.
В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами и , появляются гаметы и , и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов.
Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот.
Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом.
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
| Образуются гаметы: |
Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет.
| Образуются гаметы: |
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза).
1 — Механизм цис-фазы (некроссоверные гаметы); 2 — механизм транс-фазы (некроссоверные гаметы).
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности:
- гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
- каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
- гены расположены в хромосомах в определенной линейной последовательности;
- гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
- сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
- каждый вид имеет характерный только для него набор хромосом — кариотип.
Биология. 11 класс
Как уже отмечалось, Г. Мендель, установивший важнейшие закономерности наследования, выдвинул предположение о существовании особых наследственных факторов, контролирующих признаки живых организмов.
Однако материальная природа этих факторов, впоследствии названных генами, долгое время оставалась невыясненной. Лишь в начале ХХ в., после переоткрытия законов Г. Менделя, цитологи обратили внимание на связь гипотетических наследственных факторов с поведением хромосом при мейозе и оплодотворении.
В 1902—1903 гг. немецкий эмбриолог Т. Бовери и американский цитолог У. Саттон (фамилии приведены не для запоминания) независимо друг от друга пришли к выводу, что именно хромосомы и являются носителями менделевских факторов.
Однако ученые не смогли предоставить четких доказательств своей правоты, поэтому их предположение в течение нескольких лет так и оставалось «хромосомной гипотезой».
Хромосомная теория наследственности. Сцепленное наследование. Убедительные доказательства того, что гены располагаются в хромосомах, были получены в 1910 г. американским генетиком, будущим лауреатом Нобелевской премии (1933 г.) Т. Морганом.
Многочисленные эксперименты Моргана и его сотрудников привели к ряду важнейших открытий, которые легли в основу хромосомной теории наследственности.
Одно из положений этой теории можно сформулировать следующим образом: гены расположены в хромосомах в линейном порядке и занимают определенные участки — локусы, причем аллельные гены находятся в одинаковых локусах гомологичных хромосом.
Закон независимого наследования (третий закон Менделя) справедлив только в том случае, если неаллельные гены находятся в разных парах хромосом. Однако количество генов у живых организмов значительно больше числа хромосом. Например, у человека около 25 тыс.
генов, а количество хромосом — 23 пары (2n = 46), у плодовой мушки дрозофилы приблизительно 14 тыс. генов и всего 4 пары хромосом (2n = 8). Следовательно, каждая хромосома содержит множество генов.
Будут ли гены, локализованные в одной хромосоме, наследоваться независимо? Очевидно, что нет.
Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Такое совместное наследование генов Т. Морган назвал сцепленным наследованием (в отличие от независимого).
Гомологичные хромосомы каждой пары содержат гены, контролирующие одни и те же признаки, поэтому количество групп сцепления равно числу пар хромосом. Например, у человека 23 группы сцепления, а у дрозофилы — 4.
Вам известно, что при независимом наследовании дигетерозиготная особь, например , образует четыре типа гамет в равном соотношении, т. е. по 25 %: , , и . Это обусловлено тем, что неаллельные гены находятся в разных парах хромосом. Если же они расположены в гомологичных хромосомах, следовало бы ожидать, что дигетерозигота будет производить лишь два типа гамет: 50 % и 50 % . Обратите внимание на то, что сцепленные гены записываются в одну хромосому.
Однако Т. Морган обнаружил, что в большинстве случаев дигетерозиготные особи образуют не два типа гамет, а четыре. Кроме ожидаемых и , формируются также гаметы с новыми комбинациями генов: и , но в меньшем процентном соотношении, чем при независимом наследовании. Рассмотрим один из опытов, в котором Т. Морган изучал наследование сцепленных генов у дрозофилы.
*Дрозофила обыкновенная, или плодовая, — вид насекомых отряда Двукрылые (рис. 34.1). Особи, как правило, имеют желто-коричневое брюшко с темными поперечными кольцами и глаза красного цвета. В природе эти мушки питаются опавшими фруктами, соком растений, гниющими растительными остатками.
В XX в. дрозофила стала одним из самых распространенных объектов исследований в области генетики и биологии индивидуального развития. Удобство ее использования обусловлено рядом причин.
Так, дрозофилы имеют множество отчетливо различимых альтернативных признаков, небольшие размеры (обычно около 2—2,5 мм), легко разводятся в лабораторных условиях и обладают высокой плодовитостью — от каждого скрещивания можно получить до нескольких сотен потомков.
Важно также и то, что время развития особи от откладки яйца до выхода половозрелой мушки из куколки при 25 °C занимает всего 10 дней.*
Чистую линию дрозофил, имеющих серое тело и нормальные (длинные) крылья, скрестили с чистой линией, особи которой имели черное тело и зачаточные крылья (рис. 34.2).
Полученные гибриды первого поколения в соответствии с первым законом Менделя были единообразными — серыми с нормально развитыми крыльями.
Следовательно, у дрозофилы серое тело (А) полностью доминирует над черным (а), а нормальные крылья (В) — над зачаточными (b). Все гибриды первого поколения — дигетерозиготы.
Затем было проведено анализирующее скрещивание (рис. 34.3). Дигетерозиготную самку из гибридного поколения скрестили с рецессивным дигомозиготным самцом (черное тело и зачаточные крылья).
В потомстве было получено по 41,5 % особей с серым телом, нормальными крыльями и черным телом, зачаточными крыльями, а также по 8,5 % мух с серым телом, зачаточными крыльями и черным телом, нормальными крыльями.
Если бы гены, определяющие цвет тела и развитие крыльев, находились в разных парах хромосом, соотношение фенотипических классов было бы равным — по 25 %. Но этого не наблюдалось, значит, гены находятся в гомологичных хромосомах и наследуются сцепленно.
Несмотря на сцепление генов, самка производила не два, а четыре типа гамет. Однако гамет с исходными сочетаниями сцепленных генов формировалось намного больше ( и вместе составили 83 %), чем с новыми их комбинациями (сумма и равна 17 %).
Было выяснено, что причиной появления хромосом с новыми комбинациями родительских генов является кроссинговер. Вы знаете, что этот процесс происходит в профазе I мейоза и представляет собой обмен соответствующими участками между гомологичными хромосомами.
Таким образом, кроссинговер препятствует полному (абсолютному) сцеплению генов. Гаметы, которые образуются в результате кроссинговера, и особи, которые развиваются при участии таких гамет, называются кроссоверными или рекомбинантными.
В рассмотренном эксперименте гаметы и являлись кроссоверными, а гаметы и — некроссоверными (см. рис. 34.3).
*Т. Морган подвергал анализирующему скрещиванию не только дигетерозиготных самок дрозофилы, но и самцов, т. е. проводил реципрóкные скрещивания.
Так называют пару скрещиваний, в одном из которых организмы с исследуемыми признаками (или признаком) используются в качестве материнских, а в другом — в качестве отцовских.
Оказалось, что реципрокные анализирующие скрещивания дигетерозиготных особей дрозофилы дают разные результаты.
В потомстве дигетерозиготного самца наблюдалось лишь два фенотипических класса (рис. 34.4). Половину потомков составляли особи с серым телом и нормальными крыльями, вторую половину — особи с черным телом и зачаточными крыльями.
Это свидетельствовало об отсутствии кроссинговера между сцепленными генами или, иначе говоря, о полном (абсолютном) сцеплении генов. Выяснилось, что у самцов дрозофилы в ходе мейоза действительно не происходит кроссинговер между гомологичными хромосомами.
Впоследствии подобное явление было обнаружено у самцов некоторых других видов двукрылых, а также у самок тутового шелкопряда. Однако полное сцепление генов наблюдается в природе очень редко, как исключение из общего правила.*
Кроссинговер между сцепленными генами происходит с определенной вероятностью (частотой). Для расчета частоты кроссинговера (rf, от англ. recombination frequency — частота рекомбинации) можно пользоваться следующей формулой:
.
Таким образом, между генами А и В, контролирующими цвет тела и длину крыльев дрозофилы, кроссинговер происходит с частотой: rfAB = 17 %.
Дальнейшие исследования, проведенные Т. Морганом и сотрудниками его лаборатории, показали, что частота кроссинговера пропорциональна расстоянию между генами, расположенными в одной хромосоме.
Чем больше расстояние между сцепленными генами, тем чаще между ними происходит кроссинговер. И наоборот, чем ближе друг к другу расположены гены, тем меньше частота кроссинговера между ними.
Чем объясняется эта закономерность?
В профазе I мейоза при конъюгации гомологичных хромосом взаимный обмен участками между хроматидами осуществляется произвольно, в любой их точке. Рассмотрим рисунок 34.5.
Гены А и В (или а и b) находятся сравнительно близко друг к другу. Вероятность того, что обмен произойдет именно на участке, разделяющем эти гены, невелика.
Гены А и D (или а и d) располагаются на значительном расстоянии друг от друга. Поэтому вероятность того, что хроматиды совершат обмен в какой-либо точке между этими генами, намного выше.
Значит, чем больше расстояние между генами, тем чаще они разделяются при кроссинговере.
Таким образом, частота кроссинговера позволяет судить о расстоянии между генами. В честь Т. Моргана единица измерения расстояния между генами получила название морганида или, что то же самое, сантиморган (сМ).
Морганида (сантиморган, сМ) — это генетическое расстояние, на котором кроссинговер происходит с вероятностью 1 %.
Кроссинговер играет очень важную биологическую роль. В результате этого процесса у потомков возникают новые комбинации родительских генов, что повышает генетическое разнообразие организмов и расширяет возможности их адаптации к различным условиям окружающей среды.