Биология и медицина
Появление наземных, или высших, растений ознаменовало начало новой эры в жизни нашей планеты. Освоение растениями суши сопровождалось появлением новых, наземных, форм животных; сопряженная эволюция растений и животных привела к колоссальному разнообразию жизни на земле, изменила ее облик.
Первые достоверные наземные растения, известные только по спорам , датируются началом силурийского периода . Из верхнесилурийских и нижнедевонских отложений по сохранившимся макроостаткам или по отпечаткам органов описаны наземные растения. Эти первые известные нам высшие растения объединены в группу риниофитов .
Несмотря на анатомическую и морфологическую простоту строения, это были уже типичные наземные растения. Об этом свидетельствует наличие кутинизированной эпидермы с устьицами, развитой водопроводящей системы, состоящей из трахеид , и наличие многоклеточных спорангиев с кутинизированными спорами.
Следовательно, можно предположить, что процесс освоения суши растениями начался значительно раньше – в кембрии или в ордовике .
Предпосылок для появления наземных растений было, по-видимому, несколько. Во-первых, независимый ход эволюции растительного мира подготовил появление новых, более совершенных форм.
Во-вторых, за счет фотосинтеза морских водорослей в атмосфере земли произошло увеличение количества кислорода; к началу силурийского периода оно достигло такой концентрации, при которой оказалась возможной жизнь на суше.
В-третьих, в начале палеозойской эры на обширных территориях земли происходили крупнейшие горообразовательные процессы, в результате которых возникли Скандинавские горы, горы Тянь-Шань, Саяны. Это вызвало обмеление многих морей и постепенное появление суши на месте бывших мелких водоемов.
Если раньше водоросли , населявшие литоральную зону, только в отдельные кратковременные периоды жизни оказывались вне воды, то по мере обмеления морей они переходили к более длительному пребыванию на суше. Это, очевидно, сопровождалось массовой гибелью водорослей; выживали лишь те немногие растения, которые смогли противостоять новым условиям жизни.
В ходе длительного эволюционного процесса возникали новые виды, постепенно формировавшие типичные наземные растения.
К сожалению, промежуточные формы палеонтологическая летопись не сохранила. Новая воздушно-наземная среда обитания оказалась чрезвычайно противоречивой, принципиально отличной от первоначальной водной. Прежде всего она характеризовалась повышенной солнечной радиацией, дефицитом влаги и сложными контрастами двухфазной воздушно-наземной среды.
Вполне вероятно допустить, что у некоторых переходных форм в процессе обмена веществ мог вырабатываться кутин , который откладывался, на поверхности растений. Это было первым этапом формирования эпидермы . Чрезмерное выделение кутина неминуемо приводило к гибели растений, так как сплошная пленка кутина препятствовала газообмену.
Только те растения, у которых выделялось умеренное количество кутина, смогли сформировать сложную специализированную ткань – эпидерму с устьицами, способную и защищать растение от высыхания, и осуществлять газообмен. Таким образом, важнейшей тканью наземных растений, без которой невозможно освоение суши, следует считать эпидерму.
Однако возникновение эпидермы лишало наземные растения возможности поглощать воду всей поверхностью, как это происходит у водорослей.
У самых первых наземных растений, имевших еще небольшие размеры, поглощение воды осуществлялось с помощью ризоидов – одноклеточных или многоклеточных однорядных нитей.
Однако по мере увеличения размеров тела происходил процесс формирования сложных специализированных органов – корней с корневыми волосками .
По-видимому, образование корней, начавшееся с верхнедевонского периода , в разных систематических группах растений происходило разными путями. Активное поглощение воды ризоидами и корнями стимулировало возникновение и совершенствование водопроводящей ткани – ксилемы .
У нижнедевонских растений ксилема состояла только из кольчатых и спиральных трахеид . Начиная с верхнего девона прослеживается тенденция к “оживлению” ксилемы за счет развития древесной паренхимы , которая способствовала более активному проведению воды.
Выход растений на сушу сопровождался улучшением их освещения, что активизировало процесс фотосинтеза . Это приводило к увеличению количества ассимилятов, а следовательно, к увеличению объема растений, что вызвало необходимость их морфологического расчленения.
Существует несколько взглядов на исходную форму первых наземных растений. Одни авторы считают первичной формой пластинчатую – талломную , другие – напротив, радиальную. Есть третья точка зрения, согласно которой первые наземные растения выводятся из гетеротрихальных зеленых водорослей типа хетофоровых .
Их стелющиеся части тела дали начало талломным формам, а восходящие – радиальным, т.е. талломные и радиальные структуры возникали одновременно и развивались параллельными путями.
Пластинчатые талломы оказались малоперспективными в биологическом отношении, так как они очень быстро опутали бы всю поверхность земли тонким слоем, вызвав обостренную конкуренцию за свет. Восходящие структуры, напротив, получили дальнейшее развитие и сформировали радиальные ветвящиеся осевые органы.
Вертикальное положение тела растения оказалось возможным только при условии возникновения механических тканей . (Лигнификация клеток явилась, по- видимому, следствием избытка углеводов в ходе усиленного фотосинтеза.)
Независимо от формы тела во всех группах наземных растений очень рано (с середины девона ) начала проявляться тенденция к образованию плоских боковых фотосинтезирующих органов – листьев . Подобно корням листья возникали разными путями, т.е. в разных систематических группах они имеют разное происхождение.
(Своеобразие происхождения листьев нашло отражение в терминологии; так, все листья мохообразных иногда называют филлидами , листья плауновидных – микрофиллами, или филлоидами , листья папоротникообразных – макрофиллами, или вайями . Однако эти термины не всегда раскрывают специфику листьев разных групп растений.
)
Большая поверхность листьев в сочетании с наиболее совершенной, зернистой формой хлоропластов способствовала увеличению эффективности процессов фотосинтеза , т.е. накоплению органического вещества.
Быстрое и равномерное распределение пластических веществ по всему телу растения оказалось возможным только при наличии совершенной проводящей ткани – флоэмы , которая обнаруживается уже у нижнедевонских риниофитов .
Таким образом, у высших растений произошло формирование важнейших тканей – эпидермы , ксилемы , флоэмы , механической ткани и важнейших органов – листостебельного побега и корня .
По-латыни побег – кормос (cormos) , поэтому многие авторы называют высшие растения кормофитами. Однако А.Л.
Тахтаджян справедливо отрицает универсальность этого названия, так как среди высших растений есть талломные мохообразные , а первые наземные растения – риниофиты еще не имели сформированных побегов.
Появившись в нижнем палеозое , высшие споровые растения стали быстро распространяться по поверхности земли, осваивая новые территории. Этому в значительной степени способствовало отсутствие в тот период конкуренции.
Палеозойская эра характеризовалась интенсивными видообразовательными процессами, что привело к расцвету папоротникообразных , членистых , плауновидных .
В мезозойскую эру господство в растительном мире переходит к голосеменным растениям , а в сменившую ее кайнозойскую эру расцвета достигают покрытосеменные растения .
К сожалению общее дерево растений пока не представлено в данной базе данных.
Общее дерево высших растений ( Тахтаджан, 1987 ):
Ссылки:
- ВАРИАНТЫ ФИЛОГЕНИЙ И ДАТИРОВОК
- ВЫСШИЕ РАСТЕНИЯ: ОБЩАЯ ХАРАКТЕРИСТИКА
Источник: http://medbiol.ru/medbiol/botanica/001fe62f.htm
Эволюция растений
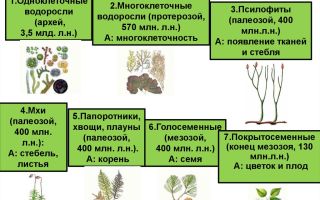
Эволюция царства растений началась с архейской эры (около 3500 млн. лет назад). В этот период на Земле отмечено появление синезеленых водорослей. Эти водоросли принадлежат к группе цианобактерий, поскольку в их клетках отсутствует оформленные ядра.
Таким образом, их можно отнести к прокариотам (доядерным организмам). Среди синезеленых водорослей были одно- и многоклеточные организмы, имеющие возможность осуществлять фотосинтез.
Благодаря процессу фотосинтеза, в атмосферу нашей планеты начал поступать кислород, необходимый для жизнедеятельности аэробов.
Позднее в протерозойской эре (около 2600 млн. лет назад) Землей завладели красные и зеленые водоросли. Их господство распространилось и на палеозойскую эру (примерно 570 млн. лет назад). Только в поздний палеозой (силурийский период) отмечена жизнедеятельность на планете древнейших высших растений – риниофитов, или псилофитов.
У этих растений были побеги, но отсутствовали корни и листья. Размножение риниофитов происходило спорами. Они обитали на суше или частично в воде.В существовании нашей планеты новая эра началась с появления высших, или наземных растений. Около 400- 360 млн.
лет назад в девонском периоде палеозойской эры на фоне преобладания на Земле риниофитов и водорослей появились первые папоротникообразные (папоротники, хвощи, плауны) и моховидные растения. Они относятся к высшим споровым растениям. Благодаря распространению растений на суше, появились и новые наземные виды животных.
Сочетанное изменение в ходе эволюции форм растений и животных обусловило огромнейшее разнообразие жизни на Земле. Облик планеты изменился коренным образом. Прикрепленный образ жизни растения на суше привел к появлению расчленения растения на корень, стебель и лист, а также к возникновению опорных тканей и сосудистой проводящей системы.
Самые первые наземные растения были маленьких размеров. Они поглощали воду посредством ризоидов, как сохранившиеся по сей день на Земле мхи. В цикле их развития преобладало гаплоидное поколение (гаметофит). Постепенно появлялись более крупные формы растений – папоротникообразные, у которых образовались сложные специализированные органы – корни с корневыми волосками.
В цикле развития этих растений на первый план выступает диплоидная фаза – спорофит, являющийся непосредственно самим растением, тогда как гаметофит – это заросток, который выглядит как клубенек у хвощей и плаунов и как небольшая пластинка в форме сердца у папоротников. Так осуществлялся постепенный переход от гаплоидного поколения к более совершенному – диплоидному.
В палеозойскую эру папоротникообразные были громадными растениями, господствовавшими на суше. Однако для их размножения была необходима вода, что ограничивало территорию их существования местностями с повышенной влажностью.
В каменноугольном периоде, который длился с 360 до 280 млн. лет назад, доказано появление на нашей планете семенных папоротников, которые в дальнейшем стали родоначальниками всех голосеменных растений.
В это время полностью исчезают риниофиты из-за невозможности конкурировать с более развитыми растениями.
А господствовашие тогда огромнейшие древовидные папоротникообразные после отмирания образовали залежи каменного угля.
В следующем пермском периоде палеозойской эры на Земле появились древние голосеменные растения. Древовидные папоротникообразные постепенно вымирают, а им на смену приходят семенные и травянистые папоротники, завладевая сушей.
Особенностью голосеменных растений является то, что размножение их осуществляется семенами, не имеющими защиты в виде стенок плода, так как цветков и плодов эти растения не образуют. Половое размножение этих растений осуществлялось независимо от капельно-водной среды.
А их появление в ходе эволюционных метаморфоз было обусловлено перепадами влажности и температуры и изменением рельефа Земли вследствие поднятия суши, то есть появления горных массивов.
Мезозойская эра наступила около 240 млн. лет назад. В триасовом периоде мезозоя появились современные голосеменные, а в юрском периоде зародились первые покрытосеменные растения. Но господствующие позиции на планете сохранились за голосеменными растениями. Это эра вымирания древних папоротникообразных, не выдерживающих естественного отбора.
В процессе появления покрытосеменных растений произошла череда ароморфозов. Во-первых, сформировался цветок – трансформированный побег, приспособленный для образования спор и гамет. Опыление, оплодотворение и образование зародыша и плода происходило непосредственно в цветке.
Во-вторых, для лучшей защиты и распространения семена покрытосеменных растений были окружены околоплодником. Для этих растений характерно половое размножение. К покрытосеменным относятся травянистые растения, деревья и кустарники. Разнообразные видоизменения вегетативных органов (корня, стебля, листа) отмечаются у разных видов растений.
Эволюционные изменения покрытосеменных растений происходили за относительно короткий срок, поэтому для них свойственна высокая эволюционная пластичность. Огромное значение в протекании эволюционных преобразований сыграли насекомые-опылители.
Покрытосеменные растения более продуктивно осваивают окружающую среду и завоевывают новые территории, благодаря своим особенностям строения и способности формировать сложные многоярусные сообщества.
В кайнозойской эре, наступившей приблизительно 70 млн. лет назад, на нашей планете стали господствовать существующие и в настоящее время покрытосеменные и голосеменные растения, тогда как высшие споровые растения регрессировали.
Сейчас на Земле произрастают более 350 видов растений, среди них встречаются цветковые, мохообразные растения, папоротники, водоросли.
Источник: http://beaplanet.ru/evolyuciya_rasteniy
Происхождение растений
Растения и животные произошли от одного примитивного предка в ходе длительной эволюции. Случилось это несколько миллиардов лет назад. Растениям, в отличие от животных, свойственно наличие хлорофилла, придающего им зеленую окраску.
Благодаря хлорофиллу растения, по способу питания, являются автотрофными, то есть способными питаться неорганическими веществами и за счет энергии солнца создавать из них нужную им органику (белки, углеводы, сахар, крахмал и прочее).
Происхождение растений и их развитие шло по пути постоянного увеличения уровней сложности от простейших одноклеточных организмов до современных цветковых растений.
Сине-зеленые водоросли или цианобактерии являются одними из первых и самых примитивных обитателей нашей планеты, отмеченных еще 3.5 млрд. лет назад. 3.3 млрд. лет назад появляются нитчатые формы. Сообщество цианобактерий способно создавать строматолиты — минеральные структуры, хорошо известные из докембрия и образующиеся и сегодня в определенных условиях (Австралия).
Появившиеся несколько позже (3 млрд. лет назад) водоросли были представлены уже и многоклеточными формами. С этого же возраста известны геологические следы процесса фотосинтеза. 1.25 млрд. лет известны красные водоросли, уже сходные с современными формами. 1.2 млрд. лет назад появляются зеленые водоросли, которые широко распространены в биотах венда и кембрия.
Произошли от зеленых жгутиковых организмов. В венде отмечены отпечатки бурых водорослей. В силуре возникают высокоорганизованные формы водорослей. 1200 млн. лет назад водоросли уже распространились по суше, обитая во влажных местах.
Однако возникновение развитых наземных форм не могло произойти до тех пор, пока озоновый слой не достиг достаточной мощности, чтобы защитить наземных обитателей от ультрафиолетового излучения.
От водорослей хлорофитов, а возможно от бурых, независимо от высших растений, произошли мхи, которые еще не имели сосудов, но уже заселяли сушу.
Их остатки найдены в карбоне, но понятно, что группа развилась намного раньше, будучи среди первых наземных растений. Споры, похожие на споры печеночных мхов, найдены в ордовике (Llanvirn).
Однако дальнейшего развития эта группа не получила. Существует также версия происхождения мхов от риниофитов.
Силур. Первые наземные растения
Следующим этапом в эволюции растений стало возникновение трахеофитов — сосудистых растений (450 млн. лет назад), которые сначала размножались спорами (споры известны с позднего ордовика), а затем и семенами. Первые наземные растения, очевидно, появились в форме крошечных растений, напоминающих печеночные мхи, в среднем ордовике.
Они еще не имели сосудистых тканей, что строго ограничивало их размеры и жестко привязывало к влажным наземным условиям. С ордовика до конца девона существовали первые наземные растения — риниофиты. Они имели кутикулу, защищающие их от высыхания, ползучее корневище и вертикальные дихотомические стебли не более 20 см высотой, но настоящих листьев еще не было.
С позднего силура они широко распространились по полузатопленным берегам мелководных лагун и рек.
В позднем силуре от псилофитовых форм происходят плауны, которые в карбоне достигают своего максимального развития. Их представители, лепидодендроны и сигиллярии были уже высокими деревьями до 30–40 м. Росли, в основном, в болотистых районах. Ветвление было еще дихотомическим, а листья имели шиловидную форму. Все ранние растения еще были споровыми.
Девонский период стал временем развития основных групп растений. В начале или середине девона из тримерофитовых развиваются членистостебельные растения — хвощи.
Они расселялись уже по берегам озер и рек, в том числе и на песчаных почвах. Их расцвет также пришелся на каменноугольный период, когда возникли уже настоящие леса.
В начале мезозойской эры стали угасать и еще до начала палеогена остался только один существующий сейчас род.
Разнообразные папоротниковые и покрытосемянное растение – глоссоптерис
В позднем девоне произошли папоротникообразные растения, широко распространившиеся в мезозойскую эру, когда существовали крупные древовидные формы, сейчас уже вымершие. Многие современные семейства появились в меловом периоде. Оледенения вытеснили их в южные широты.
Со среднего девона по ранний карбон существовали праголосемянные растения, имевшие кустистые или древовидные формы, но размножавшиеся еще при помощи спор. В перми растения расселились по внутриматериковым и горным районам.
В позднем девоне от праголосемянных возникают голосеменные растения. Первыми их них произошли семенные папоротники, существовавшие с девона до юрского периода. Позже появились кордаиты (ранний карбон), саговники (пермь), гинкговые (пермь) и хвойные (поздний карбон). В мезозойскую эру они господствовали в растительном покрове на всей Земле.
На границе юры и мела появляются покрытосеменные или цветковые растения, которые в наше время являются самыми многочисленными. Сначала развилась группа двудольных, которая уже в раннем мелу дала начало однодольным растениям, являющимся, в основном, травянистыми. Вот так происходило происхождение растений.
Источник: http://dinoera.com/bakterii-prosteyshie-i-rasteniya/proishozhdenie-rasteniy
Эволюция растений
Cлайд 1
Выполнила: Студентка v курса эколого- биологического факультета Лунина В. А.
Cлайд 2
Эволюция растений была изучена по ископаемым остаткам древних растений и с помощью тщательного сравнения строения разных современных растений. От жгутиконосцев произошли низшие Одноклеточные – водоросли. В процессе эволюции одноклеточные водоросли, пройдя через этап колониальности, перешли к многоклеточным. Благодаря многоклеточности появилась специализация.
Cлайд 3
Cлайд 4
Развитие водорослей характерно для архея, протерозоя, кембрика, ордовика. Зеленые водоросли возникли раньше других растений на Земле. 580 млн.
лет понадобилось растениям, чтобы перейти от водного к наземному образу жизни! Завоевание суши было длительным и трудным процессом, стоившим многих жертв растениям. Водоросли появились примерно 1 млрд. лет назад.
А первые наземные растения появились только 420 млн. лет назад.
Cлайд 5
Во-первых, в воде менее сильно действует гравитация на тело, поэтому ему нужно было иметь какую-то определенную форму тела, и в итоге, приобрести опору. Опора появилась в виде механических тканей. Растения в ходе эволюции постепенно потеряли подвижность и перешли к прикрепленному образу жизни.
Они в течение всей жизни остаются на одном месте, а передвижение связано либо с размножением (гаметы, пыльца, споры), либо с постепенным нарастанием органов (например, корней в почве). Необходимые для фотосинтеза диоксид углерода, свет и вода находятся в двух средах – воздушной и почвенной.
Поэтому нужно, чтобы часть растения находилась в почве, а часть – в воздушной среде, то есть одновременно они должны присутствовать в двух средах. Кроме того, чтобы проводить воду из почвы вверх, должна была появиться транспортная система.
Для того, чтобы выйти на сушу, растениям надо было решить ряд проблем.
Cлайд 6
Следующая задача заключалась в защите от обезвоживания. Сухопутная среда способствует обезвоживанию, поэтому растения должны были прибрести приспособления для добывания и сохранения воды.
Первые растения были покрыты толстым слоем воскоподобного вещества кутина, то есть они защищались от обезвоживания кутикулой. Затем в процессе эволюции появилась ткань эпидермис.
Для фотосинтеза и дыхания нужно, чтобы газообмен происходил не с раствором (как в случае с водорослями), а с воздушной средой. Для этого у растений существуют такие образования – устьица.
Нежные половые клетки должны быть защищены, а мужские гаметы – подвижные сперматозоиды – могут двигаться только в воде. В процессе эволюции произошел переход к образованию неподвижных мужских гамет – спермиев и доставке их к яйцеклетке с помощью пыльцевой трубки. Для того, чтобы выйти на сушу, растениям надо было решить ряд проблем:
Cлайд 7
Происхождение наземных растений Считается, что все наземные растения, существующие сейчас, произошли от риниофитов (тип споровых растений, составленный самыми примитивными сосудистыми формами).
Сейчас наземные растения представлены моховидными, псилотовидными, плауновидными, хвощевидными, папоротниковидными, голосеменными и цветковыми растениями.
Предполагается, что предками всех существующих растений были водоросли, в частности, зеленые.
Cлайд 8
Группы наземных растений и родственные связи между ними
Cлайд 9
Cлайд 10
2 пути образования листьев выросты покровных тканей (энационное происхождение) образовались из разветвленных осей, путем их дальнейшего уплощения (теломное происхождение)
Cлайд 11
Cлайд 12
Cлайд 13
Эти плоские ветки особенно хорошо представлены у папоротников. Здесь целые системы осей образовывали листья, поэтому они могут быть такими большими, разветвленными. У голосеменных и цветковых листья образовались из уплощенных конечных веточек.
Cлайд 14
Появление транспортной системы Что касается транспортной системы – она должна быть двух типов. Одна должна проводить воду от корней к листьям, а другая – проводить вещества, образовавшиеся в листьях к разным органам растений.
Ткани, по которым идет восходящий ток (то есть вода с растворенными в ней минеральными веществами) называется ксилемой. Кроме того, эта ткань еще выполняет дополнительную опорную функцию. Клетки ксилемы могут действовать только в мертвом состоянии.
Для того, чтобы вода проходила беспрепятственно, содержимое клетки отмирает, и вода поступает наверх по капиллярному типу. Вторая ткань – это флоэма. Она осуществляет проведение того, что образовалось в листьях ко всем органам, которым нужны эти вещества – это нисходящий ток.
У флоэмы клетки в живом состоянии проводят эти вещества. Эта ткань мягкая, так как стенки клеток не одревесневшие.
Cлайд 15
На рисунке ксилема окрашена красным цветом.
Cлайд 16
Размножение растений
Cлайд 17
Мхи – это растения, которые пошли по пути преобладания в жизненном цикле гаметофитного поколения. То, что мы видим в лесу – гаметофиты. На их верхушке образуются мужские и женские половые органы.
Спорофит отдельно жить не может, он паразитирует на гаметофите, и представляет собой коробочку, в которой образуются споры. Все остальные наземные растения пошли по другому пути – пути преобладания в жизненном цикле спорофита.
Спорофит получил расчленение на органы, он устроен довольно сложно. Гаметофит представляет собой маленькое растеньице. Размножение растений
Cлайд 18
Споровые растения образуют мужские половые клетки – сперматозоиды, окончание – зоид означает, что они движутся в воде, и это приводит к тому, что эти растения могут жить только там , где есть достаточное количество воды для размножения.
В геологическом прошлом Земли процветание этих видов было тогда, когда климат был очень влажный, их расцвет приходится на каменно-угольный период, и именно они образовали залежи каменного угля, которыми мы до сих пор пользуемся.
Но вскоре климат изменился, изменились условия, эти растения стали вымирать, на смену им пришли другие растения, и эти растения должны были научиться не использовать воду для оплодотворения. Доставка мужских клеток к яйцеклеткам должна была происходить без участия воды.
Такими растениями стали семенные: голо- и покрытосеменные.
Cлайд 19
Появление семенных растений
Cлайд 20
Cлайд 21
Захват новых участков при вегетативном размножении (с помощью образования усов, корневищ или корней, на которых впоследствии образуются новые надземные побеги) Расселение с помощью диаспор Диаспо ра (от греч.
diaspeiro «рассеиваю, распространяю») — часть растения, естественным образом отделяющаяся от него и служащая для размножения и расселения. Различают вегетативные диаспоры (клубень, луковица, выводковая почка) и генеративные (споры, семена, плоды, соплодия).
Иногда диаспорой может стать и вся надземная часть растения или целое растение; это характерно для морфологической формы растений, известных как перекати-поле. Способы расселения растений
Cлайд 22
Способы распространения диаспор автохори я (от греч. autos «сам» и -хория) — распространение диаспор без участия каких-либо посредников; механохори я (от греч.
mechane «орудие» и -хория) — распространение семян в результате разбрасывания их вскрывающимися плодами (другое название — автомеханохория); барохори я (от греч. baros «тяжесть» и -хория) — опадение семян под действием собственной массы; аллохори я (от греч.
allos «другой» и -хория) — распространение диаспор при помощи внешних факторов: человека, животных, ветра, воды…
Cлайд 23
Способы расселения растений
Cлайд 24
Основные этапы эволюции растений Эра Период Начало млн. лет назад Эволюционные события Катархей не подразд. 3900 Химическая эволюция, приведшая к возникновению биополимеров. Архей не подразд. 3500— 3800 Возникновение жизни на Земле, появление первых клеток — начало биологической эволюции.
Появление анаэробных автотрофных организмов, бактерий, цианобактерий. Протерозойская (ранней жизни) Поздний протерозой 1650 Развитие водорослей, различных многоклеточных примитивных организмов. Ранний протерозой 2600 Развитие одноклеточных прокариотических и эукариотических фотосинтезирующих организмов.
Возникновение полового процесса.
Cлайд 25
Эра Период Начало млн. лет назад Эволюционные события Палеозойская (древней жизни) Кембрий 570 Жизнь сосредоточена в морях. Эволюция водорослей; развитие многоклеточных форм. Ордовик 500 Обилие морских водорослей. Предположительное появление первых наземных растений — риниофитов. Силур 435 Возникновение современных групп водорослей и грибов.
В конце периода достоверное появление первых наземных растений. Девон 410 Расцвет риниофитов, к началу позднего девона их вымирание. Появление современных типов сосудистых растений. Карбон 345 Расцвет плауновидных, хвощевидных, папоротниковидных, семенных папоротников; появление хвоцных. Пермь 285 Распространение первых групп голосеменных.
Cлайд 26
Эра Период Начало млн. лет назад Эволюционные события Мезозойская (средней жизни) Триас 240 Вымирание семенных папоротников. Развитие голосеменных. Юра 195 Появление диатомовых водорослей. Господство папоротников и голосеменных.
Мел 136 В начале периода господство голосеменных и появление покрытосеменных, которые преобладают во второй половине периода. Кайнозойская (новой эры) Палеоген 66 Расцвет диатомовых водорослей и основных групп покрытосеменных. Неоген 2,5 Преобладание покрытосеменных и хвойных, отступание лесов, увеличение площади степей.
Четвертичный 2,4 Вымирание многих видов растений, упадок древесных форм, расцвет травянистых; растительный мир приобретает современный облик.
Cлайд 27
1. Выход на сушу морских водорослей; примитивные наземные растения. 2. Бессеменные растения. Мхи, плауны, хвощи и папоротники. 3. Появление семян. Голосеменные. 4. Появление покрытосеменных, характеризующихся наличием цветов и плодов. 5. Развитие злаковых трав и позднее хлебных злаков. В результате изучения ископаемых остатков можно установить следующие основные моменты в развитии растений:
Cлайд 28
30.01.2017 скрыт30.01.2017 скрыт30.01.2017 скрыт30.01.2017 скрыт30.01.2017 скрыт30.01.2017 скрыт30.01.2017 скрыт30.01.2017 скрыт30.01.2017 скрыт30.01.2017 скрыт30.01.2017 скрыт30.01.2017 скрыт
504625046350464504655046650467504685046950471504725047350474
Скачать эту презентацию
Источник: http://bigslide.ru/biologiya/11314-evolyuciya-rasteniy.html
Презентация на тему “Высшие растения “
- Слайд 1
- Высшие растения подразделяются на две группы: высшие споровые и семенные растения.
- По сравнению с низшими растениями число видов высших очень велико и достигает по данным разных систематиков от 300 000 до 500 000.
- Слайд 2
- К высшим споровым растениям относят отделы:
- моховидные, псилофитовидные, псилотовидные, плауновидные, хвощевидные и папоротниковидные.
- Семенные растения включают отделы: голосеменные и покрытосеменные.
- Слайд 3
- Высшие растения произошли скорее всего от водорослей.
- Чередование поколений у высших споровых также аналогично чередованию поколений многих водорослей.
- Половые мужские клетки высших споровых имеют жгутики, так же как и гаметы или зооспоры водорослей. Имеется сходство в строении и функциях хлоропластов.
- Предками высших растений, вероятно, являлись зеленые пресноводные нитчатые или гетеротрихальные водоросли.
- Слайд 5
- Эволюция жизненного цикла высших растений шла в двух противоположных направлениях.
- У моховидных она была направлена в сторону возрастания самостоятельности гаметофита и его постепенного морфологического расчленения, потере самостоятельности и редукции спорофита – его морфологического упрощения.
- Остальные группы пошли по пути прогрессивного развития спорофита
- Слайд 6
- После выхода растений на сушу, который, вероятно, произошел в девоне, они попали в совершенно иную экологическую обстановку, отличную от условий жизни в воде и вынуждены были выработать специальные структуры, органы, ткани для сухопутной жизни.
- Слайд 7
- Все органы спорофитов высших растений покрыты особым защитным слоем эпидермой, или кожицей.
- Для транспирации и газообмена с внешней средой в эпидерме в процессе эволюции возникли особые устройства –устьица, состоящие из 2-х замыкающих клеток
- Слайд 8
- Для высших растений характерно наличие многоклеточныхполовых органов (гаметангиев) и многоклеточных спорангиев органов бесполого размножения.
- Мужские половые органы называются антеридии, а женские архегонии. В антеридиях развиваются мужские гаметы (сперматозоиды или спермии), а в архегониях женские гаметы(яйцеклетки).
- Слайд 9
- С усилением половой дифференциации количество мужских гамет в гаметангиях возрастало, а количество женских, наоборот, уменьшалось.
- Таким путем возник в конце концов архегоний с одной большой и неподвижной яйцеклеткой.
- Слайд 10
- Чем ниже уровень эволюционного развития высших растений, тем в большей степени их половой процесс зависит от воды.
- У споровых растений оплодотворение яйцеклетки подвижными сперматозоидами происходит только при наличии воды, да и развитие самого гаметофита и даже спорофита нуждается в повышенной влажности.
- Слайд 11
- На спорофите формируются органы бесполого размножения спорангии со спорами.
- Перед образованием спор в спорангиях у высших растений всегда происходит мейозредукционное двухступенчатое деление, при котором из одной диплоидной материнской клетки образуются 4 гаплоидные споры.
- Споры имеют защитные оболочки – экзину и интину.
- Слайд 12
- Гаплоидные споры прорастают в гаметофит – заросток
- На гаметофитах в гаметангиях формируются гаметы: сперматозоиды и яйцеклетки
- В результате оплодотворения, т.е. слияния мужской гаметы с женской, образуется зигота с двойным набором хромосом (отцовским и материнским).
- Зигота при прорастании дает начало многоклеточному зародышу, из которого развивается спорофит – диплоидное поколение.
- Слайд 13
- Из оплодотворенной яйцеклетки развивается зародыш.
- У современных споровых он имеет ножку, посредством которой внедряется в ткань заростка (гаметофита) и получает от него питательные вещества.
- У зародыша папоротника, например, имеются зачаточный корешок,стебелек и первый лист.
- Слайд 15
- Моховидные – единственный отдел, представляющий гаплоидную линию эволюции высших растений.
- Основной жизненной формой у моховидных является гаплоидное поколение – гаметофит, выполняющий кроме своей основной функции функцию фотосинтеза.
- Спорофит- бесполое поколение – сильно редуцирован и паразитирует на гаметофите, питаясь за его счет.
- Слайд 16
ОТДЕЛ BRYOPHYTA – МОХОВИДНЫЕ
- Среди высших растений моховидные образуют обособленную группу.
- Собственно мхами систематики сейчас называют представителей листостебельных или настоящих мхов (класс Bryopsida). Отрасль систематики, изучающая мхи, называется бриологией.
- Древность моховидных подтверждается ископаемыми находками.
- В настоящее время известно примерно 23000- 27000 видов моховидных.
- Слайд 17
- антеридий
- архегоний
- зигота
- спорангий
- сперматозоид
- Вода!!!
- протонема
- гаметофит
- Слайд 18
- гаметофит мхов помимо своей основной функции – осуществлять половой процесс, выполняет и важные вегетативные функции – фотосинтез, водное и минеральное питание.
- Слайд 19
- На гаметофите формируются половые органы (гаметангии) – антеридии и архегонии.
- Обычно они располагаются группами, перемежаясь с многочисленными стерильными нитями – парафизами, и окружены видоизмененными листовидными выростами.
- Слайд 20
- Архегонии представлены бутыльчатым многоклеточным образованием с узкой шейкой и брюшком, где помещается крупная яйцеклетка.
- При созревании архегония в шейке ослизняются особые канальцевые и шейковые клетки и по образующемуся каналу при наличии капельно-жидкой воды сперматозоиды могут проникнуть к яйцеклетке.
- Слияние гамет и дальнейшее развитие зиготы происходит внутри архегония.
- Слайд 22
Зигота прямо в архегонии прорастает в спорогон, на котором формируются органы бесполого размножения со спорами.
У высших споровых всегда перед образованием спор происходит редукционное деление (мейоз), поэтомуспоры – гаплоидны
- Слайд 23
- Спора прорастает в многоклеточное ветвистое или пластинчатое тело – протонему, или предросток, похожее на водоросль.
- Протонема – юношеская стадия развития гаметофита, характерная только для моховидных
- Слайд 25
- На протонеме формируются новые многочисленные гаметофиты
- Протонема продолжает ветвиться, на ней закладываются все новые гаметофиты
- У некоторых мхов протонема является основной жизненной формой
- Слайд 26
- антеридий
- архегоний
- зигота
- спорангий
- сперматозоид
- Вода!!!
- протонема
- гаметофит
- Слайд 27
- Схистостега перистая
- Буксбаумия
- Слайд 28
- Таким образом, у моховидных наблюдается чередование, или смена поколений: полового поколения (гаметофита), представленного зеленым растением и бесполым (спорогоном) – развивающимся и живущим прямо на гаметофите.
- Слайд 29
- Коробочка – основная часть спорогона. Она имеет самую разнообразную форму, а у некоторых мхов -сложную морфологическую дифференциацию
- Слайд 30
На Земле произрастают около 25 000 видов моховидных.
Этот отдел традиционно разделяют на три класса (отдела):
- 1. Антоцеротовые (Anthocerotopsida),
- 2. Печеночные(Marchantiopsida) и
- 3. Листостебельные (Bryopsida) мхи.
- Слайд 31
- В классе насчитывается около 300 видов.
- Центральное место принадлежит роду Anthoceros(около 200 видов) со стручковидной коробочкой.
- Слайд 33
- КЛАСС MARCHANTIOPSIDA , ИЛИ HEPATICOPSIDA – ПЕЧЕНОЧНИКИ, ИЛИ ПЕЧЕНОЧНЫЕ МХИ
- Печеночники (около 10 000 видов) поражают разнообразием форм гаметофитов, а спорофит имеет однотипное строение. У большинства протонема развита слабо и недолговечна. Среди печеночников есть талломные формы, но большинство имеет листостебельные побеги со спинно-брюшным расположением листьев.
- Слайд 34
- Антеридии печеночников часто имеют округлую форму и сидят на небольшой ножке. У слоевищных форм они находятся в особых вместилищах – антеридиальных камерах, которые погружены в ткань слоевища или особых “подставок” на ножках (Marchantia – рис. 5).
- Слайд 35
- Слайд 36
- Выводковые колбочки
- Выводковые тельца на листьях
- Слайд 37
- Пор. Marchantiales – Маршанциевые:
- Это талломные напочвенные мхи
- Marchantia polymorpha и Conocephalum
- Водный мохRiccia
- Слайд 38
- Порядок Marchantiales – Маршанциевые
- Маршанциевые – обитатели влажных лесов или водные мхи. Наибольшего разнообразия они достигают в тропических и субтропических регионах, где среди них много эпифитов. В нашей флоре довольно часто можно встретить маршанцию многообразную(Marchantia polymorpha),
- Слайд 42
- Riccia glauca
- Riccia fluitans
- Слайд 44
- Порядок Metzgeriales – Метцгериевые:
- Pellia, Blasia
- Порядок Jungermanniales – Юнгерманние-вые:
- Plagiochila, Radula, Ptilidium
- Слайд 50
- Листостебельные бриевые мхи включают наибольшее число (около 15000) видов и имеют первостепенное значение среди других мхов по участию в растительных сообществах различных зон.
- Слайд 52
- У ортотропных мхов гаметангии и спорогоны развиваются на вершине стебля, таким образом мхи получаются верхоплодными.
- Плагиотропные мхи, наоборот, бокоплодны
- Слайд 53
- Основная ткань
- Центральный пучок
- гиалодерма
- Слайд 54
- крышечка
- урночка
- спорангии
- колонка
- Коробочка политрихума
Спорогон листостебельных мхов имеет сложное строение и состоит из коробочки, стопы и ножки. В коробочке выделяют крышечку, колечко, урночку, шейку и апофизу. В коробочке споры формируются вокруг особой колонки
- Слайд 56
- Апофиза – расширенная часть коробочки
- Она часто окрашена
- Слайд 57
- Для постепенного и своевременного рассеивания спор по краю урночки развит зубчатый гигроскопичный перистом
- Слайд 60
- Все листья однослойные, без жилки и состоят из клеток двух типов – узких хлорофиллоносных и широких бесцветных мертвых со спиральными утолщениями и порами (гиалиновых, или водоносных). Благодаря гиалиновым клеткам сфагновые мхи очень гигроскопичны и обладают огромной влагоемкостью
- Слайд 62
- Слайд 63
- В подклассе только один порядок Sphagnales с одним семейством сфагновых и одним родом сфагнум (Sphagnum), в котором насчитывается свыше 300 видов.
- Во флоре России известно около 40 видов сфагновых мхов.
- Слайд 64
- Андреевые мхи являются мелкими, жесткими и хрупкими черно-бурыми мхами, растущими в виде подушечек на камнях и скалах.
- В подклассе всего один порядок андреевые (Andreaeales) и одно семейство с родами андрея (Andreaea) и неуролома(Neuroloma).
- Слайд 67
- Зрелая коробочка напоминает китайский фонарик и раскрывается 4-8 продольными щелями-створками,
- Слайд 68
Порядки:
- политриховые (Polytrichales)
- бриевые (Bryales)
- изобриевые (Isobryales)
- гипнобриевые (Hypnobryales)
- фунариевые (Funariales)
- гриммиевые (Grimmiales)
- Слайд 69
- Слайд 70
- Слайд 72
- Долгомошно-сфагновое болото
- Слайд 74
- Слайд 75
- Слайд 76
- Слайд 77
- Слайд 81
- моховидные произошли непосредственно от водорослей (так называемая “водорослевая” теория происхождения
- Моховидные произошли от одной из групп древнейших высших споровых в результате общей редукции спорофита и прогрессивного развития гаметофита (“риниофитовая” теория происхождения
Посмотреть все слайды
Источник: https://pptcloud.ru/biologiya/gaploidnaya-liniya-i-mohovidnye
Общая характеристика высших растений, их классификация. Основные направления эволюции высших растений
Высшие растения составляют подцарство растительного мира. Существует предположение, что они произошли от каких-то древних групп зеленых водорослей.
Для такой гипотезы имеются веские основания: 1) у водорослей и высших растений главный фото-синтезирующий пигмент — хлорофилл а и каротиноиды; 2) основной запасной углевод — крахмал, который откладывается в хлоропластах, а не в цитоплазме, как у других фотосинтезирующих эукариот; 3) целлюлоза является важнейшим компонентом их клеточной стенки; 4) у водорослей и некоторых высших растений (мхи) в матриксе хлоропласта имеются особые включения — пиреноиды; 5) при клеточном делении у растений и некоторых водорослей образуются фрагмопласт — внутриклеточная пластинка, зачаток клеточной стенки.
Высшие растения появились на суше примерно 430 млн. лет назад в виде небольших по размеру и примитивных по строению риниофитов, или псилофитов. В дальнейшем их эволюция неразрывно связана с постепенным завоеванием суши.
Оказавшись в совершенно иной наземновоздушной среде, они постепенно адаптировались к необычной обстановке и на протяжении многих миллионов лет дали громадное разнообразие наземных растений различной величины и сложности строения.
Одним из ключевых событий раннего этапа выхода растений на сушу было появление слор с прочными оболочками, позволяющими переносить засушливые условия и распространяться ветром.
В процессе дальнейшего приспособления к наземным условиям существования у высших растений сформировались вегетативные органы — корень, стебель и лист, как результат разделения функций между различными участками тела.
Корни обеспечивают закрепление растений в субстрате и водно-минеральное питание, листья — фотосинтез, стебли — транспорт веществ (восходящий и нисходящий токи).
Развитие эффективной проводящей системы тканей, состоящей из ксилемы и флоэмы, решило проблему передвижения воды и органических веществ у сосудистых растений при выходе их на сушу.
От высыхания эти растения обрели защиту в виде покровной ткани — эпидермы или перидермы.
Развитие эпидермы привело к возникновению устьиц — высокоспециализированных образований, регулирующих газообмен и испарение воды растением.
Далее у высших растений произошло усиление механической прочности путем утолщения клеточной стенки и пропитывания ее лигнином, придающим жесткость ее целлюлозному остову.
Параллельно шла эволюция и органов полового (антеридии и архегонии) и бесполого (спорангии) размножения. Из одноклеточных, характерных для подавляющего большинства водорослей, эти органы становятся многоклеточными, и стенки их надежно защищают развивающиеся гаметы и споры от высыхания.
В жизненном цикле высших наземных растений наблюдается закономерное чередование полового и бесполого поколений.
Гаплоидное поколение называется гаметофитом, поскольку оно способно к половому размножению и образует гаметы. Гаметы формируются в антеридиях и архегониях.
В результате оплодотворения образуется зигота, из которой вырастает диплоидный спорофит. Он способен к бесполому размножению с образованием гаплоидных спор. Последние дают начало гаметофит-ному поколению.
Одно из этих двух поколений всегда преобладает над другим, и на его долю приходится большая часть жизненного цикла высших растений.
В отличие от других высших растений, в жизненном цикле моховидных преобладает гаметофит —небольшое, преимущественно листостебельное растение, которое осуществляет функцию фотосинтеза, водоснабжения и минерального питания.
Спорофит у них развивается из оплодотворенной яйцеклетки внутри архегония и постоянно связан с гаметофитом не только морфологически, но и физиологически (в смысле питания), т. е. низведен до степени органа растения, выполняющего только функцию спорообразования. Для размножения мхов необходима вода, иначе сперматозоиды не смогут подплыть к архегони-ям.
Кроме того, только в достаточно влажной среде у них лопаются антеридии и высвобождаются сперматозоиды. Поэтому не случайно большинство моховидных обитает в сырых и затененных местах.
Развитие жизненного цикла моховидных по пути возрастания самостоятельности гаметофита и морфологического упрощения (с потерей самостоятельности) спорофита привело к эволюционному тупику.
В эволюции высших сосудистых растений происходит постепенная редукция (уменьшение и упрощение) гаметофита и преобладание в жизненном цикле спорофита.
Так, у плауновидных, хвощевидных и папоротниковидных гаметофит представлен маленьким (от нескольких миллиметров до 3 см) заростком, не расчлененным на органы, живущим несколько недель (у плаунов — несколько лет) независимо от спорофита.
На заростках в антеридиях развиваются сперматозоиды, которые, плавая в каплях воды, достигают архегония и сливаются с яйцеклеткой. Благодаря крошечным размерам гаметофитов оплодотворение у хвощей, плаунов и папоротников может происходить даже при ничтожно малых количествах воды в виде капелек росы, тумана и Др.
У голо- и покрытосеменных растений гаметофит полностью утратил способность к самостоятельному образу жизни, и все его развитие протекает на спорофите внутри макроспорангия (или семязачатка).
У голосеменных женский гаметофит — многоклеточный гаплоидный эндосперм с двумя (у сосны) или несколькими (у других голосеменных) архегониями; у покрытосеменных он редуцирован обычно до семи клеток, архегониев не имеет и называется зародышевым мешком. В последнем образуются яйцевой аппарат, состоящий из яйцеклетки и двух клеток-синергид, вторичного диплоидного ядра и клеток-антипод.
Мужской гаметофит семенных растений развивается из микроспоры и представляет собой пыльцевое зерно (пыльцу), прорастающее в пыльцевую трубку с образованием двух спермиев.
При этом впервые в эволюции растений процесс оплодотворения становится независимым от наличия капельно-жидкой среды: спермин доставляются к яйцеклеткам пыльцевой трубкой, что является важнейшим приспособлением к наземному образу жизни.
Источник: https://megaobuchalka.ru/10/3457.html
Происхождение и систематика высших растений
Высшие растения, вероятно, произошли от каких-либо водорослей. Об этом свидетельствует то, что в геологической истории растительного мира высшим растениям предшествовали водоросли.
В пользу этого предположения свидетельствуют и следующие факты: сходство наиболее древней вымершей группы высших растений – риниофитов – с водорослями, очень сходный характер их ветвления; сходство в чередовании поколений высших растений и многих водорослей; наличие жгутиков и способность к самостоятельному плаванию у мужских половых клеток многих высших растений; сходство в строении и функции хлоропластов.
Предполагают, что высшие растения произошли скорее всего от зеленых водорослей, пресноводных или солоноватоводных. Они имели многоклеточные гаметангии, изоморфное чередование поколений в цикле развития.
Первыми наземными растениями, найденными в ископаемом состоянии, были риниофиты (риния, хорнея, хорнеофитон, спорогонитес, псилофит и др.).
После выхода на сушу высшие растения развивались в двух основных направлениях и образовали две большие эволюционные ветви – гаплоидную и диплоидную.
Гаплоидная ветвь эволюции высших растений представлена отделом моховидные (Bryophyta). В цикле развития мхов преобладает гаметофит, половое поколение (само растение), а спорофит, бесполое поколение, редуцирован и представлен спорогоном в виде коробочки на ножке.
Вторая эволюционная ветвь высших растений представлена всеми остальными высшими растениями.
Спорофит в наземных условиях оказался более жизнеспособным и адаптированным к разнообразным экологическим условиям. Эта группа растений более успешно завоевывала сушу.
В настоящее время высшие растения насчитывают свыше 300 000 видов. Они господствуют на Земле, населяют ее от арктических территорий до экватора, от влажных тропиков до сухих пустынь. Они образуют различные типы растительности – леса, луга, болота, заполняют водоемы. Многие из них достигают гигантских размеров.
Систематика высших растений – это раздел ботаники, который разрабатывает естественную классификацию высших растений на основе изучения и выделения таксономических единиц, устанавливает родственные связи между ними в их историческом развитии.
Важнейшими понятиями систематики являются таксономические (систематические) категории и таксоны.
Под таксономическими категориями подразумевают определенные ранги или уровни в иерархической классификации, полученные в результате последовательного подразделения абстрактного множества на подмножества.
Согласно правилам ботанической номенклатуры основными таксономическими категориями считаются: вид (species), род (genus), семейство (familia), порядок (ordo), класс (classis), отдел (devisio), царство (regnum). При необходимости могут использоваться и промежуточные категории, например, подвид (subspecies), родрод (subgenus), подсемейство (subfamilia), надпорядок (superordo), надцарство (superregnum).
Для видов, начиная с 1753 г. – даты выхода в свет книги К. Линнея “Виды растений” – приняты биноминальные названия, состоящие из двух латинских слов. Первое обозначает род, к которому относится данный вид, второе – видовой эпитет: например ольха клейкая –Alnus glutinosa.
Для семейств растений принято окончание – aceae, для порядков – ales, для подклассов – idae, для классов – psida, для отделов – phyta. В основу стандартного униноминального названия кладется название какого-либо рода, включаемого в это семейство, порядок, класс и т.д.
Современная наука об органическом мире делит живые организмы на два надцарства: доядерные организмы (Procariota) и ядерные организмы (Eucariota). Надцарство доядерных организмов представлено одним царством – дробянки (Mychota) с двумя подцарствами: бактерии (Bacteriobionta) и цианотеи, или сине-зеленые водоросли (Cyanobionta).
Надцарство ядерных организмов включает три царства: животные (Animalia), грибы (Mycetalia, Fungi, или Mycota) и растения (Vegetabilia, или Plantae).
Царство животных делится на два подцарства: простейшие животные (Protozoa) и многоклеточные животные (Metazoa).
Царство грибов подразделяется на два подцарства: низшие грибы (Myxobionta) и высшие грибы (Mycobionta).
Царство растений включает три подцарства: багрянки (Rhodobionta), настоящие водоросли (Phycobionta) и высшие растения (Embryobionta).
| · Низшие растения o Тип Сине-зеленые водоросли o Класс Разножгутиковые водоросли o Класс Диатомовые водоросли o Отдел Зеленые водоросли o Класс Бурые водоросли o Отдел Красные водоросли o Царство Грибы o Лишайники | · Высшие растения o Надотдел Мохообразные o Отдел Плауновидные o Отдел Папоротниковидные o Класс Хвощёвые o Класс Папоротниковые o Тип Голосеменные o Тип Покрытосеменные |
Дата добавления: 2016-05-30; просмотров: 2447;
Источник: https://poznayka.org/s2609t1.html