Много людей использует термин ДНК. Но статей, нормально описывающих, как она работает почти нет (понятных не биологам). Я уже описывал в общих чертах устройство клетки и самые основы ее энергетических процессов. Теперь перейдем к ДНК. ДНК хранит информацию. Это знают все. Но вот как она это делает? Начнем с того, где она в клетке хранится. Примерно 98% хранится в ядре. Остальное в митохондриях и хлоропластах (в этих ребятах протекает фотосинтез). ДНК — это огромный полимер, состоящий из мономерных звеньев. Выглядит примерно так.
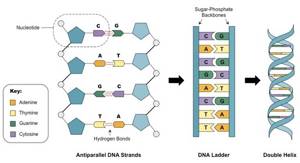
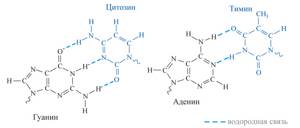
Что мы тут видим? Во-первых ДНК — двухцепочечная молекула. Почему это так важно — чуть позже. Далее мы видим синие пятиугольники. Это молекулы дезоксирибозы (такой сахар, чуть меньше глюкозы. От рибозы отличается отсутствием одной OH группы, что придает стабильности молекуле ДНК, в отличие от РНК, в которой используется рибоза. Дальше, для простоты опущу приставку дезокси и буду просто говорить рибоза, да простят нас щепетильные товарищи). Маленькие кружкИ — остатки фосфорной кислоты. Ну и собственно есть азотистые основания. Всего их 5, но в ДНК в основном встречаются 4. Это Аденин, Гуанин, Тимин и Цитозин. То есть, есть рибоза с которой связано азотистое основание. Вместе они образуют так называемые нуклеозиды, которые связываются друг с другом с помощью остатков фосфорной кислоты. Таким образом мы получаем длинную цепь, состоящую из мономеров. Теперь посмотрите на увеличенную левую цепь. Видите C и G соединены тремя пунктирными линиями, а T и A двумя. Что это значит? Да, ДНК состоит из двух цепей, но что удерживает их вместе? Есть такая штука, как водородная связь. Выглядит примерно так. На атомы кислорода (O) и азота (N) формируется частичный отрицательный заряд, а на водороде (H) — положительный. Это приводит к формированию слабых связей.
Связи действительно очень слабые. Их энергия может быть в 200 раз ниже энергии ковалентных связей (образуются за счет перекрытия пары электронных облаков, например связь в молекуле CO2). Однако таких связей много. В каждой нашей клетке ДНК цепи связаны почти 16 миллиардами слабых связей, не мало, согласны? Но вернемся к числу связей между основаниями. Цитозин и Гуанин связаны тремя связями, а Аденин и Тимин — двумя. Это приводит к тому, что Г и Ц связанны куда прочнее, чем А и Т. Некоторым организмам нужна особая стабильность связей ДНК, например живущим при высоких температурах. При нагревании ДНК содержащая больше ГЦ пар более стабильна. Так что хочешь жить в гейзере — имей много ГЦ пар. Хотя последние исследования говорят, что явной связи между GC составом (% ГЦ пар от всех пар) и температурой обитания нет. Стоит сказать, что варьирует он сильно. Так у Candidatus Carsonella ruddii PV (внутриклеточный эндосимбионт) он примерно 16%, у нас с вами почти 41%, а у Anaeromyxobacter K (бактерия вполне себе средних размеров) достигает 75%. Тут вы можете видеть связь GC состава с размером генома бактерий. Mb — миллион пар нуклеотидов. Показатель довольно вариативный. Его, кстати, часто юзают как фичу при обучении различного рода классификаторов. Сам недавно писал классификатор для распознания патогенов на основе сырых данных секвенирования и оказалось, что GC состав даже по одному риду вполне себе можно использовать. Пока не забыл. Почему важно, что ДНК двухцепочечная? На основе одной цепи можно восстановить другую. Если в одной цепи поврежден кусок напротив последовательности Аденин-Аденин-Цитозин, то мы точно знаем, что до повреждения там был Тимин-Тимин-Гуанин. Таким образом наличие второй цепи позволяет надежней хранить информацию. Круто! Теперь вернемся к самой молекуле ДНК. Это цепочка из 4х типов звеньев. Однако насколько длинная? У Candidatus Carsonella ruddii PV уже упомянутого выше всего 160 000 нуклеотидов. У нас с вами 3.2 миллиарда (в гаплоидной клетке, то есть с одним набором хромосом. У большинства наших клеток их два). Кажется много, да? На самом деле нет. У одноклеточной амебы (Amoeba dubia) он примерно 670 миллиардов пар нуклеотидов. Кажется что это бесконечно длинная цепочка, поэтому давайте переведем размер в любимые нам метры. Если все наши хромосомы (их 46, не забываем; 23 по две копии на каждую) развернуть и вытянуть в одну линию, получится примерно 2х метровая цепочка. ДНК одной амебы хватит, чтоб опоясать футбольный стадион. Но к чему я веду? Ядро, в котором ДНК хранится не очень большое. У нас оно в среднем диаметром в 6 мкм. Не очень то много, если хочешь свернуть 2х метровую нить, пусть и очень тонкую. Причем нужно не просто запихать нить в ядро. Нужно свернуть таким образом, чтобы в любой момент можно было обеспечить доступ к любому ее участку. Задача сложная. И с ней успешно справляются специализированные белки. Они создают ряд спиралей и петель, которые обеспечивают все более и более высокие уровни упаковки и не до допускают спутывания ДНК в гордиев узел. Давайте поговорим о том, как она упаковывается. Сразу скажу, упаковывается она очень по разному. Но если откинуть экзотику, то остается два способа. Первый характерен для бактерий, второй для эукариот (или иначе ядерных).
 Связи действительно очень слабые. Их энергия может быть в 200 раз ниже энергии ковалентных связей (образуются за счет перекрытия пары электронных облаков, например связь в молекуле CO2). Однако таких связей много. В каждой нашей клетке ДНК цепи связаны почти 16 миллиардами слабых связей, не мало, согласны? Но вернемся к числу связей между основаниями. Цитозин и Гуанин связаны тремя связями, а Аденин и Тимин — двумя. Это приводит к тому, что Г и Ц связанны куда прочнее, чем А и Т. Некоторым организмам нужна особая стабильность связей ДНК, например живущим при высоких температурах. При нагревании ДНК содержащая больше ГЦ пар более стабильна. Так что хочешь жить в гейзере — имей много ГЦ пар. Хотя последние исследования говорят, что явной связи между GC составом (% ГЦ пар от всех пар) и температурой обитания нет. Стоит сказать, что варьирует он сильно. Так у Candidatus Carsonella ruddii PV (внутриклеточный эндосимбионт) он примерно 16%, у нас с вами почти 41%, а у Anaeromyxobacter K (бактерия вполне себе средних размеров) достигает 75%. Тут вы можете видеть связь GC состава с размером генома бактерий. Mb — миллион пар нуклеотидов. Показатель довольно вариативный. Его, кстати, часто юзают как фичу при обучении различного рода классификаторов. Сам недавно писал классификатор для распознания патогенов на основе сырых данных секвенирования и оказалось, что GC состав даже по одному риду вполне себе можно использовать.
Связи действительно очень слабые. Их энергия может быть в 200 раз ниже энергии ковалентных связей (образуются за счет перекрытия пары электронных облаков, например связь в молекуле CO2). Однако таких связей много. В каждой нашей клетке ДНК цепи связаны почти 16 миллиардами слабых связей, не мало, согласны? Но вернемся к числу связей между основаниями. Цитозин и Гуанин связаны тремя связями, а Аденин и Тимин — двумя. Это приводит к тому, что Г и Ц связанны куда прочнее, чем А и Т. Некоторым организмам нужна особая стабильность связей ДНК, например живущим при высоких температурах. При нагревании ДНК содержащая больше ГЦ пар более стабильна. Так что хочешь жить в гейзере — имей много ГЦ пар. Хотя последние исследования говорят, что явной связи между GC составом (% ГЦ пар от всех пар) и температурой обитания нет. Стоит сказать, что варьирует он сильно. Так у Candidatus Carsonella ruddii PV (внутриклеточный эндосимбионт) он примерно 16%, у нас с вами почти 41%, а у Anaeromyxobacter K (бактерия вполне себе средних размеров) достигает 75%. Тут вы можете видеть связь GC состава с размером генома бактерий. Mb — миллион пар нуклеотидов. Показатель довольно вариативный. Его, кстати, часто юзают как фичу при обучении различного рода классификаторов. Сам недавно писал классификатор для распознания патогенов на основе сырых данных секвенирования и оказалось, что GC состав даже по одному риду вполне себе можно использовать. 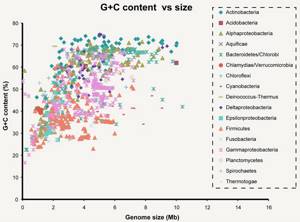 Пока не забыл. Почему важно, что ДНК двухцепочечная? На основе одной цепи можно восстановить другую. Если в одной цепи поврежден кусок напротив последовательности Аденин-Аденин-Цитозин, то мы точно знаем, что до повреждения там был Тимин-Тимин-Гуанин. Таким образом наличие второй цепи позволяет надежней хранить информацию. Круто! Теперь вернемся к самой молекуле ДНК. Это цепочка из 4х типов звеньев. Однако насколько длинная? У Candidatus Carsonella ruddii PV уже упомянутого выше всего 160 000 нуклеотидов. У нас с вами 3.2 миллиарда (в гаплоидной клетке, то есть с одним набором хромосом. У большинства наших клеток их два). Кажется много, да? На самом деле нет. У одноклеточной амебы (Amoeba dubia) он примерно 670 миллиардов пар нуклеотидов. Кажется что это бесконечно длинная цепочка, поэтому давайте переведем размер в любимые нам метры. Если все наши хромосомы (их 46, не забываем; 23 по две копии на каждую) развернуть и вытянуть в одну линию, получится примерно 2х метровая цепочка. ДНК одной амебы хватит, чтоб опоясать футбольный стадион. Но к чему я веду? Ядро, в котором ДНК хранится не очень большое. У нас оно в среднем диаметром в 6 мкм. Не очень то много, если хочешь свернуть 2х метровую нить, пусть и очень тонкую. Причем нужно не просто запихать нить в ядро. Нужно свернуть таким образом, чтобы в любой момент можно было обеспечить доступ к любому ее участку. Задача сложная. И с ней успешно справляются специализированные белки. Они создают ряд спиралей и петель, которые обеспечивают все более и более высокие уровни упаковки и не до допускают спутывания ДНК в гордиев узел. Давайте поговорим о том, как она упаковывается. Сразу скажу, упаковывается она очень по разному. Но если откинуть экзотику, то остается два способа. Первый характерен для бактерий, второй для эукариот (или иначе ядерных).
Пока не забыл. Почему важно, что ДНК двухцепочечная? На основе одной цепи можно восстановить другую. Если в одной цепи поврежден кусок напротив последовательности Аденин-Аденин-Цитозин, то мы точно знаем, что до повреждения там был Тимин-Тимин-Гуанин. Таким образом наличие второй цепи позволяет надежней хранить информацию. Круто! Теперь вернемся к самой молекуле ДНК. Это цепочка из 4х типов звеньев. Однако насколько длинная? У Candidatus Carsonella ruddii PV уже упомянутого выше всего 160 000 нуклеотидов. У нас с вами 3.2 миллиарда (в гаплоидной клетке, то есть с одним набором хромосом. У большинства наших клеток их два). Кажется много, да? На самом деле нет. У одноклеточной амебы (Amoeba dubia) он примерно 670 миллиардов пар нуклеотидов. Кажется что это бесконечно длинная цепочка, поэтому давайте переведем размер в любимые нам метры. Если все наши хромосомы (их 46, не забываем; 23 по две копии на каждую) развернуть и вытянуть в одну линию, получится примерно 2х метровая цепочка. ДНК одной амебы хватит, чтоб опоясать футбольный стадион. Но к чему я веду? Ядро, в котором ДНК хранится не очень большое. У нас оно в среднем диаметром в 6 мкм. Не очень то много, если хочешь свернуть 2х метровую нить, пусть и очень тонкую. Причем нужно не просто запихать нить в ядро. Нужно свернуть таким образом, чтобы в любой момент можно было обеспечить доступ к любому ее участку. Задача сложная. И с ней успешно справляются специализированные белки. Они создают ряд спиралей и петель, которые обеспечивают все более и более высокие уровни упаковки и не до допускают спутывания ДНК в гордиев узел. Давайте поговорим о том, как она упаковывается. Сразу скажу, упаковывается она очень по разному. Но если откинуть экзотику, то остается два способа. Первый характерен для бактерий, второй для эукариот (или иначе ядерных). Упаковка ДНК у бактерий
Начнем с братьев наших меньших. Бактерии сами по себе обладают не очень большим геномом, в среднем от 1 до 5 миллионов пар нуклеотидов. Наиболее характерное их отличия от нас в том, что у них нет ядра и ДНК плавает в клетке. Не совсем плавает, оно частично прикреплено к клеточной мембране и тоже свернуто, но не так сильно как у нас. Второе.
Бактериальная ДНК чаще всего кольцевая. Так ее проще копировать (нет концов, которые могут потеряться при копировании и не нужно придумывать механизмы сохранения концов). Обычно такое кольцо одно, но у некоторых бактерий их может быть 2 или 3. Есть еще кольца поменьше (от пары тысяч до пары сотен тысяч остатков).Имя им плазмиды, и это вообще отдельная история.
Вернемся к упаковке ДНК. ДНК упаковывают белки-гистоны (есть еще гистоноподобные белки). ДНК это дезоксирибонуклеиновая кислота. Кислота. Это значит что она отрицательно заряжена (за счет остатков фосфорной кислоты). Поэтому белки, связывающие ее положительно заряжены. Таким образом они могут связываются с ДНК. ДНК бактерий вместе с белками ее упаковывающими формируют нуклеоид, при этом на долю ДНК приходится 80% от его массы. Выглядит это примерно так. То есть кольцевая ДНК делится на домены по 40 тысяч пар нуклеотидов. Затем происходит скручивание. Внутри доменов тоже происходит скручивания, но его степень в разных доменах отличается. В среднем степень упаковки бактериальной ДНК варьирует от сотни до тысячи раз.
Есть еще прикольное видео.
Упаковка ДНК у эукариот
Тут все куда интересней. Наше ДНК хорошо упакована и спрятана внутри ядра. И она куда эффективней упакована, нежели у бактерий. Во время митоза (деление клетки) размер 22й хромосомы составляет 2 мкм. Если ее распутать и вытянуть, она будет уже 1,5 см.
Что соответствует степени упаковки в 10 000 раз. Это около максимальная степень упаковки нашей ДНК. Во время деления нужно максимально упаковать ДНК, что бы эффективно разделить ее между дочерними клетками. В обыденной жизни степень компактизации составляет примерно 500 раз.
Со слишком упакованной ДНК сложно считывать информацию.
Есть несколько уровней упаковки ДНК эукариот
Первый — нуклеосомный уровень. 8 белков-гистонов формируют частицу на которую наматывается ДНК. Затем еще один белок ее фиксирует. Выглядит примерно так. Получаются своего рода бусы. Плотность упаковки благодаря этому возрастает в 7-10 раз. Далее нуклеосомы упаковываются в фибрилы. Немного похоже на солениод. Тут суммарная степень упаковки может достигать 60 раз. Следующий этап компактизации ДНК связан с образованием петлеобразных структур, которые называются хромомерами. Фибрила разбита на участки по 10 — 80 тысяч пар азотистых оснований. В местах разбивки находятся глобулы негистоновых белков. ДНК — связывающие белки узнают глобулы негистоновых белков и сближают их. Образуется устье петли. Средняя длина петли включает примерно 50 тысяч оснований. Эту структуру называют интерфазной хромонемой. И именно в ней наше ДНК находится большую часть времени. Уровень упаковки здесь достигает 500-1500 раз. При необходимости клетка может еще больше компактизировать генетический материал. Идет образование более крупных петель из хромомерной фибриллы. Эти петли в свою очередь образуют новые петли (петли в петли… и это не вязание). Которые в конечном счете формируют хромосому. В целом процесс упаковки можно описать примерно так. В итоге из нитей ДНК мы получаем, при делении, суперскрученные структуры, которые можно увидеть под микроскопом. Их мы и зовем хромосомами. Собственно вещество хромосом зовется хроматином. И степень его упаковки отличается в зависимости от участка хромосомы. Есть эухроматин и гетерохроматин. Эухроматин это довольно расплетенная область хроматина, в ней ДНК находится на хромомерном уровне (упаковка в 500 — 1000 раз). Здесь происходит активное считывание информации. Например, если сейчас клетка активно синтезирует белок А, то область ДНК, его кодирующая будет в состоянии эухроматина, что бы ферменты, «читающие» ДНК могли до нее добраться. Гетерохроматин же содержит ту часть ДНК, которая клетке не особо нужна сейчас. То есть ДНК максимально плотно упакована, дабы не путаться под ногами. В зависимости от потребностей клетки одни области хроматина могут частично расплетаться, в то время как другие — сплетаться. Таким образом еще и осуществляется регуляция (очень грубое приближение), ведь к скрученной области не добраться, и значит ее не прочитать.
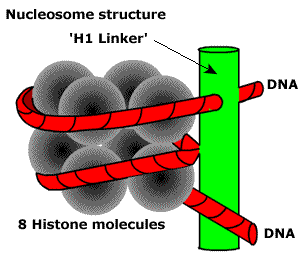
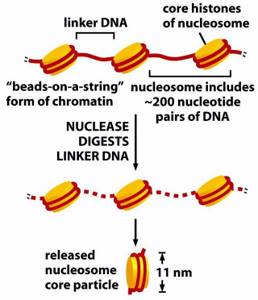 Получаются своего рода бусы. Плотность упаковки благодаря этому возрастает в 7-10 раз. Далее нуклеосомы упаковываются в фибрилы. Немного похоже на солениод. Тут суммарная степень упаковки может достигать 60 раз.
Получаются своего рода бусы. Плотность упаковки благодаря этому возрастает в 7-10 раз. Далее нуклеосомы упаковываются в фибрилы. Немного похоже на солениод. Тут суммарная степень упаковки может достигать 60 раз. 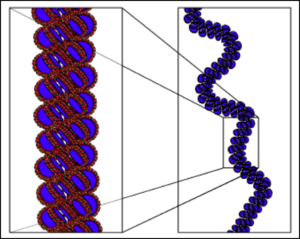 Следующий этап компактизации ДНК связан с образованием петлеобразных структур, которые называются хромомерами. Фибрила разбита на участки по 10 — 80 тысяч пар азотистых оснований. В местах разбивки находятся глобулы негистоновых белков. ДНК — связывающие белки узнают глобулы негистоновых белков и сближают их. Образуется устье петли. Средняя длина петли включает примерно 50 тысяч оснований. Эту структуру называют интерфазной хромонемой. И именно в ней наше ДНК находится большую часть времени. Уровень упаковки здесь достигает 500-1500 раз. При необходимости клетка может еще больше компактизировать генетический материал. Идет образование более крупных петель из хромомерной фибриллы. Эти петли в свою очередь образуют новые петли (петли в петли… и это не вязание). Которые в конечном счете формируют хромосому. В целом процесс упаковки можно описать примерно так.
Следующий этап компактизации ДНК связан с образованием петлеобразных структур, которые называются хромомерами. Фибрила разбита на участки по 10 — 80 тысяч пар азотистых оснований. В местах разбивки находятся глобулы негистоновых белков. ДНК — связывающие белки узнают глобулы негистоновых белков и сближают их. Образуется устье петли. Средняя длина петли включает примерно 50 тысяч оснований. Эту структуру называют интерфазной хромонемой. И именно в ней наше ДНК находится большую часть времени. Уровень упаковки здесь достигает 500-1500 раз. При необходимости клетка может еще больше компактизировать генетический материал. Идет образование более крупных петель из хромомерной фибриллы. Эти петли в свою очередь образуют новые петли (петли в петли… и это не вязание). Которые в конечном счете формируют хромосому. В целом процесс упаковки можно описать примерно так. 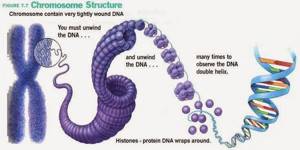 В итоге из нитей ДНК мы получаем, при делении, суперскрученные структуры, которые можно увидеть под микроскопом. Их мы и зовем хромосомами. Собственно вещество хромосом зовется хроматином. И степень его упаковки отличается в зависимости от участка хромосомы. Есть эухроматин и гетерохроматин. Эухроматин это довольно расплетенная область хроматина, в ней ДНК находится на хромомерном уровне (упаковка в 500 — 1000 раз). Здесь происходит активное считывание информации. Например, если сейчас клетка активно синтезирует белок А, то область ДНК, его кодирующая будет в состоянии эухроматина, что бы ферменты, «читающие» ДНК могли до нее добраться. Гетерохроматин же содержит ту часть ДНК, которая клетке не особо нужна сейчас. То есть ДНК максимально плотно упакована, дабы не путаться под ногами. В зависимости от потребностей клетки одни области хроматина могут частично расплетаться, в то время как другие — сплетаться. Таким образом еще и осуществляется регуляция (очень грубое приближение), ведь к скрученной области не добраться, и значит ее не прочитать.
В итоге из нитей ДНК мы получаем, при делении, суперскрученные структуры, которые можно увидеть под микроскопом. Их мы и зовем хромосомами. Собственно вещество хромосом зовется хроматином. И степень его упаковки отличается в зависимости от участка хромосомы. Есть эухроматин и гетерохроматин. Эухроматин это довольно расплетенная область хроматина, в ней ДНК находится на хромомерном уровне (упаковка в 500 — 1000 раз). Здесь происходит активное считывание информации. Например, если сейчас клетка активно синтезирует белок А, то область ДНК, его кодирующая будет в состоянии эухроматина, что бы ферменты, «читающие» ДНК могли до нее добраться. Гетерохроматин же содержит ту часть ДНК, которая клетке не особо нужна сейчас. То есть ДНК максимально плотно упакована, дабы не путаться под ногами. В зависимости от потребностей клетки одни области хроматина могут частично расплетаться, в то время как другие — сплетаться. Таким образом еще и осуществляется регуляция (очень грубое приближение), ведь к скрученной области не добраться, и значит ее не прочитать. Собственно пока это все. Мы обсудили как хранится носитель информации. Сделаем небольшую паузу и через пару дней поговорим о самом кодировании информации.
Как все закручено
Мы привыкли представлять себе ДНК в виде двойной спирали — но это лишь одно из множества ее обличий. С тех пор, как Уотсон и Крик опубликовали свою модель, в клетках человека нашли тройную и четверную спираль ДНК, а еще кресты, шпильки и другие варианты переплетения — некоторые проще нарисовать, чем описать словами.
Набросать идей
Уотсон и Крик не были единственными, кто корпел над трехмерной моделью ДНК. Они даже не были первыми. На обрывках биохимических данных можно было построить самые разные молекулярные формы, и вариантов было множество.
Условия задачи у всех были одинаковы. На начало 1953 года уже было понятно, как устроен нуклеотид:
- остаток фосфорной кислоты,
- сахар,
- одно из азотистых оснований: аденин (А), гуанин (Г), тимин (Т) или цитозин (Ц).
Еще было известно, что азотистые основания разбросаны по цепи не случайно: в любой молекуле ДНК суммарное количество аденинов и гуанинов строго равнялось количеству тиминов и цитозинов.
Кроме того, на всех рентгеновских снимках Розалинд Франклин и Рэймонда Гослинга, независимо от того, какой участок ДНК на них был запечатлен, сама нить имела одну и ту же толщину.
Это означало, что форма остается неизменной при любой последовательности нуклеотидов.
Из этих вводных Лайнус Полинг и Роберт Кори собрали свою модель — тройную спираль, ощетинившуюся со всех сторон азотистыми основаниями (фосфату и сахару биохимики отвели роль внутреннего стержня). Эта конструкция выглядела неустойчивой: было непонятно, почему отрицательно заряженные фосфатные группы в центре спирали не отталкиваются друг от друга.
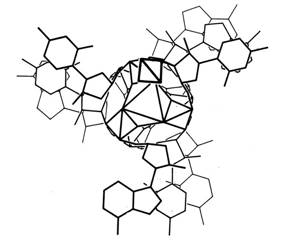
Структура ДНК по версии Полинга и Кори
Linus Pauling, Robert B. Corey / PNAS, 1953
Эту проблему решил Брюс Фрезер, вывернув конструкцию наизнанку: в его варианте три нити смотрели фосфатами наружу. Азотистые основания были обращены внутрь, однако Фрезер так и не смог объяснить, по какому принципу они соединены.
Модель Уотсона и Крика с закрученной вправо двойной спиралью оказалась самой устойчивой. Как и Фрезер, ученые расположили фосфаты снаружи, а азотистые основания — внутри. Был в этой модели и четкий принцип их противопоставления: А на одной цепи всегда соединялся с Т на другой, а Г — с Ц. Это объясняло, почему толщина конструкции стабильна — пары А-Т и Г-Ц примерно одинакового размера.
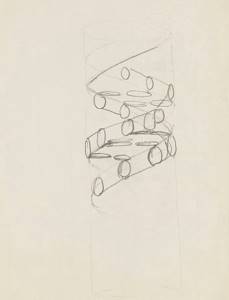
Карандашный набросок структуры ДНК, сделанный Фрэнсисом Криком
Wellcome Images / CC BY-SA 4.0
Потом были и другие попытки пересобрать ДНК в новую форму. Голландский биохимик Карст Хугстин, например, заметил, что можно соединить те же самые пары нуклеотидов другими гранями, — так спираль тоже оставалась стабильной, но получалась тоньше. Другие авторы изображали ДНК в виде спирали с чередующимися правым и левым поворотами, или даже в виде двух двойных спиралей, которые образуют единую четверку. И хотя существование Уотсон-Криковской двойной спирали с тех пор много раз подтвердилось, в XXI веке продолжают размышлять о том, какие формы принимает нить ДНК внутри клетки, где ее разглядеть намного сложнее, чем в пробирке. Правда, ни одна из альтернативных идей до сих пор не оказалась достаточно хороша, чтобы отказаться от классической правозакрученной двойной спирали.
Уотсон и Крик сделали нечто большее, чем просто разрешили споры о форме ДНК. Их модель сразу же объяснила, как эта форма работает: взаимно однозначное соответствие делает каждую нить шаблоном для другой. Имея только одну из цепей, по ней всегда можно восстановить вторую — на этот принцип опираются все современные модели передачи генетической информации.
Тем не менее, большинство «отвергнутых» идей в чем-то оказались верны. За почти 70 лет пристального разглядывания ДНК в ней удалось обнаружить практически все возможные виды соединения оснований, другие спирали и даже левый поворот.
Свернуть не туда
Уже сама по себе двойная спираль может быть устроена по-разному. Это заметила еще Розалинд Франклин, хотя и не предполагала, что перед ней спираль, да еще и двойная.
В обычных условиях, напоминающих внутриклеточные, ДНК на снимках биолога имела «рыхлую» форму, которую Франклин назвала В-ДНК.
Но если влажность в пробирке опускалась ниже 75 процентов, получалась А-ДНК, пошире и поплотнее.
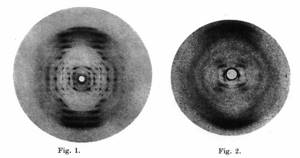
А (слева) и В (справа) формы ДНК, какими их увидела Розалинд Франклин
Rosalind Franlkin, Raymond Gosling / Acta Crystallographica, 1953
Как выяснилось потом, А-ДНК действительно закручена туже: в ней на виток спирали уходит 10 нуклеотидов, а не 11, как в В-ДНК. И расположены они не перпендикулярно оси симметрии спирали, а под углом: если в В-ДНК нуклеотиды обычно изображают горизонтальными черточками, в А-ДНК их следовало бы рисовать косыми.
Уотсон и Крик выбрали В-ДНК в качестве основы для своей модели и не прогадали. Позже оказалось, что В-вариант действительно встречается в клетке гораздо чаще, и сейчас его считают основной формой существования ДНК, а все отклонения часто обозначают общим термином «не-В ДНК».
Более того, реальная двойная спираль почти никогда не соответствует своей идиллической модели. В живых системах В-ДНК, как правило, скручена чуть сильнее, чем предсказывали Уотсон и Крик, и среднее число нуклеотидов на виток спирали в ней — не 10 и не 11, а около 10,5.
Кроме того, отдельные пары нуклеотидов постоянно отклоняются от положенной «горизонтали» (это называют «пропеллерным поворотом») поэтому спираль никогда не бывает абсолютно гладкой и ровной — то тут, то там по ее бокам торчат шероховатости: концы нуклеотидов под разными углами.
«Пропеллерный» поворот нуклеотидов в В-ДНК
James D. Watson et al. / Molecular Biology of the Gene, 2008
Позже оказалось, что витки спирали могут не только лежать туже или расслабленнее, но и вовсе закручиваться против часовой стрелки (например, влево закручена спираль башни «Эволюция» в Москва-сити, явно символизирующая нить ДНК). По странному стечению обстоятельств, именно такую ДНК увидели в 1979 году, когда появилась наконец возможность рассмотреть нуклеиновые кислоты с высоким разрешением. Это все еще была двойная спираль, но совсем другой формы: 12 нуклеотидов на виток, еще тоньше, чем В-ДНК и закрученная не вправо, а влево. Торчащие ее на поверхности фосфатные группы образовывали не плавную спираль, а зигзаг, поэтому новый вариант назвали Z-формой.
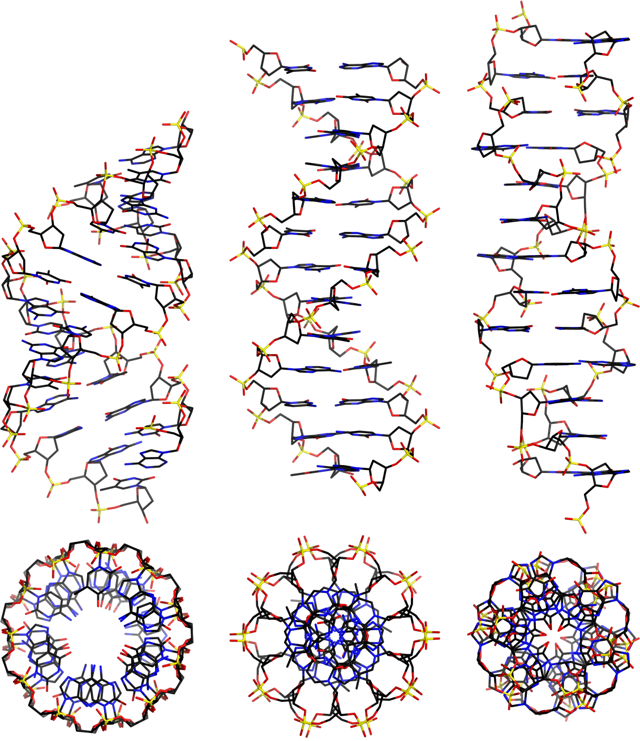
А-ДНК (слева), B-ДНК (по центру), Z-ДНК (справа)
Mauroesguerroto / wikimedia commons / CC BY-SA 4.0
Это, конечно, не означало, что Уотсон-Криковская модель неверна. Z-форму удалось получить при достаточно экзотических условиях — в растворе с высокой концентрацией солей. И в клетке она тоже получается из В-ДНК лишь при определенных обстоятельствах: например, когда «напряжение» на цепи слишком высоко и его необходимо сбросить. Напряжение появляется из-за чрезмерного скручивания: нити ДНК и так завернуты друг относительно друга, но образованная ими двойная спираль накручивается на какой-нибудь белок (например, гистон), возникает так называемая суперспирализация. Переход в Z-форму помогает сбросить напряжение и развернуть лишние витки — а это, в свою очередь, важно, чтобы с ДНК могли связываться новые белки, например, полимераза при транскрипции.
Поэтому ДНК часто принимает Z-форму при транскрипции генов. Более того, чем больше при этом Z-ДНК, тем активнее идет транскрипция. Гистоны с Z-ДНК связаться не могут, поэтому полимеразе никто не мешает заниматься своим делом. И этим, кстати говоря, активно пользуются опухолевые клетки, у которых левозакрученная спираль вовремя возникает перед нужными им генами.

Башня «Эволюция» (на переднем плане) имеет вид левозакрученной ДНК
mos.ru / CC BY-SA 4.0
Потом нашлись и другие формы двойной спирали. В зависимости от влажности, содержания солей и последовательности нуклеотидов в конкретном участке, ДНК может еще сильнее удлиняться (Е-ДНК) или сжиматься (C- и D-ДНК), включать в себя ионы металлов (М-ДНК) или вытягиваться так, что вместо азотистых оснований в центре спирали оказываются фосфатные группы (S-ДНК). А после того, как в список добавились другие типы внутриклеточной ДНК, вроде ядерной N-ДНК и рекомбинантной R-ДНК (которые, впрочем, попали в этот список не из-за своей формы, а положения в клетке или происхождения), в английском алфавите для вариантов ДНК практически закончились буквы. Тому, кто решит открыть еще какую-нибудь неканоническую форму, придется выбирать из пяти свободных: F, Q, U, V, и Y.
A-ДНК — двухцепочечная, чуть толще, чем В.
B-ДНК — та, которую построили Уотсон и Крик.
C-ДНК — двухцепочечная, 9,3 нуклеотида на виток.
D-ДНК — двухцепочечная, узкая: 8 нуклеотидов на виток, содержит много тиминов.
E-ДНК — двухцепочечная, еще уже: 15 нуклеотидов на два витка.
G-ДНК — четверная спираль с гуаниновыми тетрадами.
H-ДНК — тройная спираль.
I-ДНК — две двойные спирали, которые держатся вместе притяжением своих цитозинов.
J-ДНК — еще одна тройная спираль, которую образуют повторы АЦ.
K-ДНК — ДНК трипаносом, особенно богатая аденинами.
L-ДНК — ДНК, в основе которой лежит L-дезоксирибоза (а не D-, как обычно).
M-ДНК — В-ДНК в комплексе с двухвалентными металлами.
N-ДНК — ядерная ДНК.
O-ДНК — точка начала удвоения ДНК у бактериофага λ.
P-ДНК — тройная спираль Полинга и Кори.
R-ДНК — рекомбинатная ДНК (полученная встраиванием чужеродного фрагмента).
S-ДНК — двухцепочечная, вытянута в 1,6 раз сильнее, чем В-форма.
T-ДНК — похожа на D-форму, встречается у бактериофага Т2.
W-ДНК — синоним Z-ДНК.
X-ДНК — двухцепочечная спираль, которую образуют повторы АТ.
Z-ДНК — двухцепочечная левозакрученная.
Попасть в переплет
Помимо всевозможных форм двойной спирали и способов ее плетения, ДНК иногда распадается на отдельные нити, которые образуют в шпильки, кресты и другие двуцепочечные фигуры. Случается и так, что уже существующая двойная спираль обрастает новыми соседями.
В 1985 году выяснилось, что Полинг и Кори тридцать лет назад были правы: тройная спираль ДНК (H-ДНК) существует. Однако устроена она совсем не так, как они предполагали.
В настоящей тройной спирали две цепи соединяются стандартным, Уотсон-Криковским способом, а третья примыкает к ним сбоку, ложась в большую бороздку между цепями.
При этом азотистые основания третьей, дополнительной нити соединяются с основными парами не классическим способом, а как бы сбоку — теми самыми связями, которые предсказывал Карст Хугстин. Он тоже, в некотором роде, оказался прав.
Тройная спираль, как и многие альтернативные формы ДНК, тоже возникает в ответ на суперспирализацию цепи. Однако, в отличие от Z-формы, она не поддерживает транскрипцию, а наоборот, ей препятствует.
РНК-полимераза, которая привычно расплетает две нити перед собой, не всегда справляется с тем, чтобы разделить триплекс.
Поэтому если в гене или его регуляторных областях образуется тройная спираль, он работает хуже прочих.
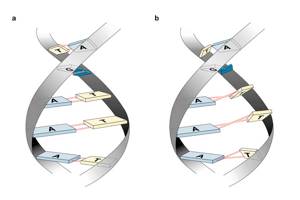
Варианты образования тройной спирали. Уотсон-Криковские пары обозначены черным, добавочный третий нуклеотид выделен цветом
Yutaro Yamagata et al. / Chemistry Europe, 2015
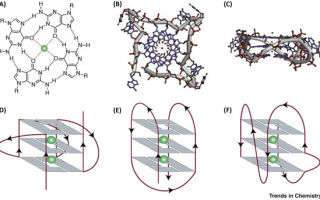
Бывает и так, что соединяются не две и не три, а сразу четыре цепи ДНК. Чтобы это произошло, в одном месте должны встретиться четыре гуаниновых нуклеотида — и неважно, находятся они на двух цепях одной нити или на четырех разных нитях, не связанных друг с другом. Каждый гуанин образует неклассическую, хугстиновскую пару с двумя соседями, а все вместе они создают квадратную гуаниновую тетраду. Если рядом с ними находятся другие гуанины, способные создать квадрат, то из них складывается стэк — стопка, которая удерживает рядом четыре цепи ДНК.
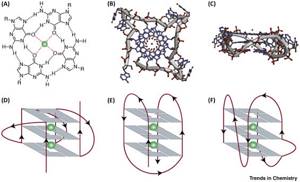
Гуаниновая тетрада (сверху) и варианты расположения цепей в квадруплексе (снизу)
Jochen Spiegel et al. / Trends in Chemistry, 2020
Все 30 лет, что прошли с момента открытия квадруплексов, количество процессов, в которых они так или иначе замешаны, растет. Известно уже больше двух сотен белков, которые могут избирательно распознавать гуаниновые тетрады — вероятно, последние выполняют роль своего рода генетической разметки, очередного способа регулировать упаковку и транскрипцию генов. Например, они часто встречаются в промоторах (регуляторных участках, с которых начинается транскрипция) разных генов. Совсем недавно ученым даже удалось отличить разные типы рака груди через наборы квадруплексов — от них, в свою очередь, зависело, какие гены в опухолевых клетках были гиперактивны.
Чем дальше мы вглядываемся в молекулу ДНК, тем больше замечаем отклонений от давно привычной модели. Двойная спираль — не единственная и не окончательная структура ДНК, а лишь одна (пусть и самая частая) из поз, которую та принимает в непрерывном танце. Повинуясь велению нуклеотидной последовательности, нить ДНК сжимается и разжимается, изгибается, закручивается и принимает бесконечное число (прекрасных) форм. Ни одна из них — не окончательная: альтернативные структуры ДНК переходят друг в друга, конкурируют с В-формой и между собой, подчиняются сигналам клеточных белков и сами направляют их работу.
Найти и возглавить
Неканонические формы ДНК, при всем своем разнообразии, не возникают в случайных местах. Устойчивость им придает определенный набор нуклеотидов в их составе, поэтому и появляются они лишь в тех участках цепи, где для них есть «удобная» последовательность.
Так, например, в ДНК есть определенные участки, которые особенно охотно сворачиваются в зигзаг.
Это места, где чередуются пары Г-Ц: после левого поворота в них каждый второй нуклеотид принимает «неправильную» форму, отсюда и ломаный профиль всей Z-формы.
Это означает, что последовательности, склонные принимать Z-форму, можно найти прямо в тексте — если видите ГЦГЦГЦГЦГЦГЦ, то вряд ли прогадаете. Так в одной работе, например, насчитали 391 такой участок в человеческом геноме.
Места, в которых может образоваться тройная спираль, тоже можно узнать по характерной последовательности нуклеотидов.
Третья цепь присоединяется либо по принципу комплементарности — то есть к паре Г-Ц добавляется еще один Г, образуя Г-Ц*Г — либо «к своему» — и получается Г*Г-Ц.
Поэтому часто такая конструкция возникает в тех местах ДНК, где подряд идет несколько одинаковых (например, ГГГГГ) или химически близких (АГГААГ) нуклеотидов и где они образуют палиндромные (зеркальные) повторы.
Точно также по тексту ДНК можно предсказать и появление квадруплексов. По результатам только одного секвенирования (собственно, прямого перевода ДНК в буквы), в геноме человека их нашлось более 700 тысяч.
Не все они, вероятно, встречаются in vivo — для этого соответствующем нитям ДНК нужно оказаться рядом в одной точке сложно устроенного клеточного ядра — однако это может означать, что четырехспиральным структурам отведена какая-то специфическая роль в жизни клетки.
Далеко не всегда образование альтернативных форм ДНК идет клетке на пользу: большинство из них куда менее прочны, чем обычная В-ДНК, и гораздо чаще рвутся.
Поэтому последовательности, которые склонны образовывать не-В формы, становятся участками генетической нестабильности и повышенного мутагенеза. Одни исследователи видят в этом двигатель эволюции — если такие участки появляются в генах, связанных с развитием организма.
Другие же винят альтернативные формы ДНК во всех видах болезней, связанных со случайными мутациями и перестановками в геноме — от опухолей до шизофрении и аутизма.
Получается, что ДНК содержит не только информацию о строении клеточных белков и РНК, но и о том, какие формы эта информация может принимать, помимо Уотсон-Криковского стандарта. А уже от этих форм, в свою очередь, зависит то, что с этой информацией произойдет: сможет ли клетка ее реализовать или ген, будет вечно молчать, а то и вовсе сломается, породив какие-то дополнительные мутации.
Вероятно, мы научимся однажды вмешиваться в этот процесс — можно было бы, например, построить цепь нуклеотидов, которая имитировала бы третью цепь в спирали и «подсунуть» ее в нужное время в нужном месте, чтобы заблокировать работу какого-нибудь нежелательного гена в клетке.
Были даже более смелые предложения — использовать тройную спираль для прицельного редактирования генома: ввести в клетку нуклеотид, который сможет образовать с целевым участком ДНК тройную спираль и побудить систему репарации заменить этот участок на «здоровый» вариант с другой хромосомы.
А пока мы этому только учимся, остается признать структуру ДНК еще одним видом информации — помимо генетической (нуклеотидного «текста») и эпигенетической (доступности генов для считывания) — который несет в себе наш геном. И нам еще предстоит научиться с ним работать, влияя через форму на содержание, или наоборот.
Полина Лосева
Структура хроматина. Упаковка ДНК
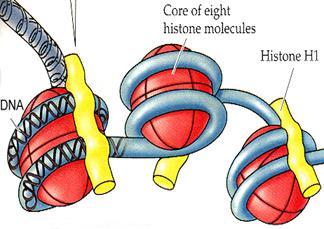
Первый уровень упаковки нитей ДНК- нуклеосомный– связан с участием в этом процессе специальных ядерных белков – гистонов.Во всех эукариотических клетках выделено пять фракций гистонов (Н1, Н2А, Н2B, H3, H4).
Группа из 8 молекул гистонов (по две молекулы каждого из гистонов Н2А, H2B, H3,H4), образуют глобулу, вокруг которой по спирали закручивается участок ДНК размером в 146 пар нуклеотидов. Эти глобулы «бусинки» были открыты в 1974 году и названы нуклеосомами.
Нуклеосомный уровень укладки хроматина позволяет «сжать» молекулу ДНК примерно в 6-7 раз.
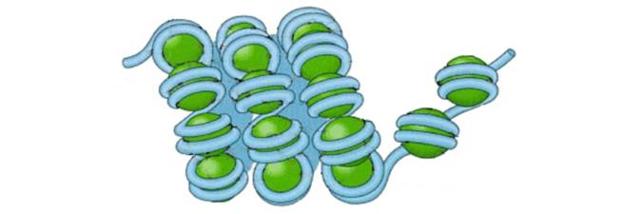
Второй уровень упаковки нитей ДНК –нуклеомерный.
несколько нуклеосом сближаются, формируют компактные группы – нуклеомеры,которые, в свою очередь образуют суперспираль.
Такая компактная нить ДНК имеет диаметр 30 нм и получила наименование – хроматиновая фибрилла. При этом происходит еще более чем 40 – кратное сжатие ДНК.
Третий уровень – петлевой или доменный. На этом уровне хроматиновые фибриллы образуют систему петель (доменов) диаметром 300 нм, в каждой из которых находится несколько генов. Это приводит к еще 600-кратному сжатию ДНК
Четвертый уровень–хромомерный Происходит переход от спирального типа укладки ДНК, состоящих из петель хроматиновых 30-нм фибрилл, к образованию компактных глобулярных структур типа хромомеров, имеющих уже размеры 0,1-0,2 мкм..Степень сжатия – несколько тысяч раз.
Пятый уровень – стадия конденсированной метафазной хромосомы
Это уровень сверхкомпактного состояния хроматина. Видны только в период митоза. Неактивны – считывание информации не происходит. К кинетохорам метафазных хромосом крепятся микротрубочки веретена деления.
- Нуклеосомы (первый уровень)
- Хиазмы Нуклеомер(второй уровень)
- Петлевой и хромомерный уровень упаковки ДНК.(третий и четвертый уровни) Видны петли ДНК и плотные (хромомерные) участки между ними
- (гигантская интерфазная хромосома в яйцеклетке птиц- типа «ламповой щетки») Стрелки указывают на участки перекреста (кроссинговера) гомологичных хромосом (хиазмы)
- Структура гена. Транскрипция
- Ген – структурная единица генетической информации, закодированная в определенной последовательности нуклеотидов участка ДНК.
- Генотип – совокупность гаплоидного набора всех генов данной клетки или организма.
- В дальнейших исследованиях было установлено, что:
- – ген дискретен и делим;
- – ген имеет очень сложную структуру;
- – информация записана в гене посредством универсального генетического кода;
- – в геноме имеются разные типы (группы) генов;

 Хиазмы Нуклеомер(второй уровень)
Хиазмы Нуклеомер(второй уровень)
– наименьшей мутирующей единицей гена является пара нуклеотидов – т.н. однонуклеотидные замены (SNP – single nucleotide polymorphisms).
Гены состоят из двух основных частей: регуляторной и кодирующей.
Кодирующая часть гена имеет мозаичную структуру и состоит из функционально разных участков – экзонов и интронов.
С одних – экзонов в дальнейшем считывается информация о белке, в других – интронахне закодирована информация о белке. Объем, занимаемый интронными участками гена, значительно превышает экзонные.
- Каждый структурный ген имеет свой промотор
- Промотор – это небольшой участок ДНК, расположенный перед структурной частью гена и предназначенный для связывания РНК-полимеразы и белков-регуляторов перед началом синтеза РНК. У высших эукариот в состав промотора входят многократно повторяющиеся последовательности нуклеотидов: ТАТА, ЦААТ, ГЦ
- Регуляторные участки гена эукариот, с которыми связываются факторы транскрипции, располагаются за сотни пар нуклеотидов от участка промотора активируемого гена- они получили название – энхансеры.
- Каждый ген или группа генов отделены друг от друга специальным участками ДНК, называемые спейсерами.
- Схема организации типичного гена эукариот и его регуляторные элементы:
- 1 – экзоны; 2 – интроны; 3 – промотор; 4 – РНК-полимераза II; 5 – главные факторы транскрипции; 6 – регуляторные белки; 7 – энхансеры (регуляторные участки гена); 8 – спейсерные участки; 9 – ТАТА блок – инициирующая часть промотора; стрелка – начало процесса транскрипции
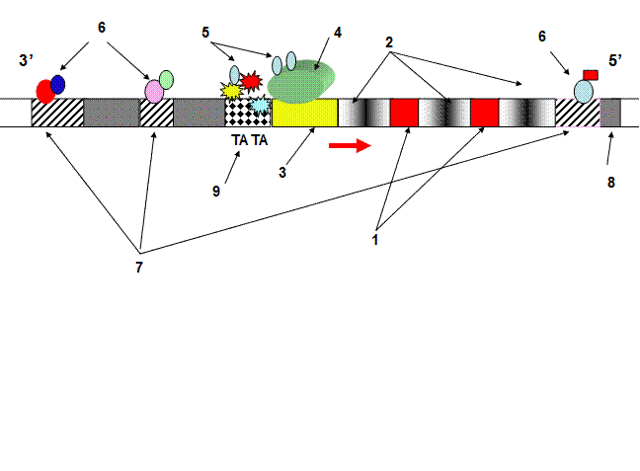 Схема организации типичного гена эукариот и его регуляторные элементы:
Схема организации типичного гена эукариот и его регуляторные элементы:В сколтехе проанализировали способы упаковки днк
Исследователи из Сколтеха и их коллеги оптимизировали анализ данных для метода изучения 3D-структуры ДНК в отдельных клетках мухи-дрозофилы. Новый подход позволяет ученым с большей уверенностью заглянуть в клетку, чтобы изучить способы упаковки ДНК, и приблизиться к пониманию основных механизмов этого крайне важного процесса.
Статья опубликована в журнале Nature Communications. Почти двухметровая нить ДНК помещается в крошечное ядро человеческой клетки благодаря тому, что хроматин, комплекс ДНК и белков, сворачивает ее в компактные, но сложные структуры.
Для изучения способов упаковки ДНК ученые во всем мире используют так называемые методы фиксации конформации хромосом (3C), и один из наиболее производительных из них — метод Hi-C.
Он позволяет обнаружить контакты ДНК всего генома с помощью высокопроизводительного секвенирования.
В этом, однако, и заключается проблема: для работы Hi-C необходимы десятки микрограммов ДНК — то есть миллионы клеток с уникальной пространственной организацией хроматина. Эту информацию приходится усреднять, чтобы получить общую картину, которая не будет учитывать особенности упаковки ДНК в отдельных клетках.
Подобно тому, как «среднестатистического человека» на самом деле не существует, традиционный метод Hi-C не может показать, какие именно из множества взаимодействий участков ДНК происходят одновременно в одной и той же клетке. Кроме того, этот «коллективный портрет» вряд ли поможет понять, какие физические процессы приводят к формированию той или иной трехмерной структуры хроматина.
«Мы видим некоторые структуры, например, так называемые топологически ассоциированные домены (ТАДы), в усредненных картах контактов ДНК, но мы не знаем, действительно ли они существуют в отдельных клетках, или же это артефакты усреднения.
Кроме того, мы знаем, что с точки зрения экспрессии генов большое разнообразие встречается даже в клетках одной и той же ткани — отсюда возникает естественный вопрос о том, насколько они разнообразны на структурном уровне», — говорит соавтор статьи Михаил Гельфанд, вице-президент Сколтеха по биомедицинским исследованиям.
Трехмерное сворачивание гаплоидной Х-хромосомы / ©www.nature.com
Чтобы разрешить эти проблемы и сделать эксперимент Hi-C более подходящим для отдельных клеток, исследователи нескольких институтов разработали метод, получивший название Hi-C одиночных клеток.
Команда Сколтеха во главе с Гельфандом и доцентом Центра наук о жизни Сколтеха Екатериной Храмеевой поставила перед собой задачу оптимизировать обработку данных для Hi-C одиночных клеток и изучить фундаментальные свойства клеток дрозофилы.
Их коллеги из Института биологии гена РАН и Московского государственного университета имени М. В. Ломоносова совместно с сотрудниками российско-французского Междисциплинарного научного центра Понселе оптимизировали метод, чтобы сделать его пригодным для экспериментов с клетками дрозофилы.
Команды начали со стандартных шагов метода Hi-C, при котором структура хроматина фиксируется химически, а ДНК разрезается и «пересобирается» так, чтобы фрагменты, которые в естественных условиях находились рядом, оказывались «сшитыми».
Но затем вместо того, чтобы использовать сразу всю ДНК, ученые амплифицировали крошечное количество ДНК из каждой клетки с помощью полимеразы бактериофага phi29.
Эта полимераза часто используется при амплификации ДНК, отчасти благодаря ее способности создавать большое количество ДНК даже по очень маленькому образцу с куда меньшим количеством ошибок, чем у других популярных полимераз.
Однако оказалось, что эта удобная ДНК-полимераза, несмотря на достаточно высокую точность копирования, все же может случайным образом «прыгать» между молекулами ДНК, создавая искусственные связи, которые алгоритм Hi-C не может отличить от настоящих взаимодействий. Поэтому исследователям пришлось придумать механизм отбраковки этих случайных «прыжков» полимеразы.
Они использовали свой новый метод на клетках дрозофилы, чтобы выяснить, имеют ли различные организмы общие фундаментальные принципы упаковки хроматина.
Предыдущие исследования на клетках млекопитающих указывали на существование ТАДов только на контактных картах, получаемых популяционного Hi-C, но не в отдельных клетках.
Однако изучение клеток дрозофилы показало, что эти домены есть и в каждой конкретной клетке.
Порядок и стохастичность в 3D-геноме дрозофилы / ©www.nature.com
Чтобы понять, какой биологический механизм отвечает за формирование этих устойчивых доменов, потребуются дополнительные исследования; пока ученые предложили две модели их возникновения.
Одна из них предполагает, что хроматин у дрозофилы организован по механизму «липкости», то есть некоторые его участки с большей вероятностью соединяются друг с другом.
Согласно другой, описывающей так называемый механизм экструзии петель, крупные белковые комплексы создают петли из нити ДНК и за счет этого упаковывают ДНК.
«Возможно, один из самых интересных вопросов заключается в том, одинаковы ли правила сворачивания хроматина у разных видов живых организмов.
С помощью метода Hi-C одиночных клеток дрозофилы мы выяснили, что в геноме этого насекомого тоже присутствуют домены, похожие на домены в клетках млекопитающих.
Однако эти структуры гораздо более упорядочены, чем у млекопитающих», — отмечает Александра Галицына, аспирантка Сколтеха и один из первых авторов статьи.
«Мы продолжим изучение архитектуры хроматина и механизмов формирования петель и ТАДов. В этой области еще много вопросов без ответов.
Мы уже знаем, что эти механизмы у некоторых организмов могут различаться, но что представляет собой эволюция сворачивания хроматина в целом? Если мы хотим понять это на достаточном уровне детализации, нам нужно заполнить пробелы, изучая структуру хроматина у странных организмов, а не только у тех, что уже хорошо исследованы. Поэтому мы уже работаем с морскими губками, дрожжами и амебами», — говорит Екатерина Храмеева.
По ее словам, группа также занимается возможной связью изменений в организации хроматина с болезнями, развитием организма и старением. «Если предположить, что архитектура хроматина тесно связана с экспрессией генов, то, ответив на эти вопросы, мы сможем разобраться в регулировании развития человеческого организма, старения и заболеваний», — отмечает Храмеева.
В исследовании приняли участие специалисты Института биологии гена Российской академии наук, Московского государственного университета имени М. В. Ломоносова, Национального центра научных исследований Франции, российско-французского Междисциплинарного научного центра Понселе и других организаций.
Маркируем гены, пакуем ДНК: ученые исследовали способы упаковки ДНК
Ученые МГУ имени М.В. Ломоносова создали метод маркировки работающих генов, основываясь на различиях активных и неактивных участков хромосом во время репликации из ДНК. Статья опубликована в журнале Current Biology.
Совместно со своими коллегами из США российские ученые исследовали способы упаковки ДНК в клеточном ядре и их изменение в процессе ее репликативного синтеза. Считается, что характер упаковки ДНК оказывает существенное влияние на работу генов и является одним из механизмов эпигенетического контроля генной экспрессии.
«Эпигенетический контроль генной экспрессии выражается в том, что, хотя все клетки организма имеют одинаковую генетическую информацию, не все гены работают в данном отдельно взятом типе клеток. Именно набор работающих генов определяет судьбу клетки.
Существуют несколько клеточных механизмов, которые помогают клетке запомнить, какие гены должны в ней работать, а какие — нет.
Это и есть эпигенетический контроль, представляющий собой память клетки о том, кто она и благодаря работе каких генов она такой является, — рассказывает один из авторов статьи, заведующий отделом электронной микроскопии НИИ физико-химической биологии имени А.Н.
Белозерского, доктор биологических наук Игорь Киреев. — Нас интересовали так называемые высшие уровни структурной организации хромосом, формирующиеся в результате серии последовательных этапов упаковки нити ДНК».
ДНК в клетке существует в виде комплекса с белками — хроматина. Начальные этапы компактизации хроматина достаточно хорошо исследованы — это нуклеосомы, белковые глобулы размером около 10 нм, состоящие из восьми молекул белков-гистонов, на которые наматывается ДНК.
Затем цепочка нуклеосом укладывается непонятным пока образом в более толстые фибриллы, хромонемы, в результате чего достигается очень высокая степень компактизации.
Так, длина максимально компактизованной митотической хромосомы в 20 000 раз меньше длины уложенной в ней ДНК.
Репликация — это процесс синтеза дочерней молекулы ДНК на матрице родительской молекулы, а транскрипция — процесс синтеза РНК с использованием ДНК в качестве матрицы.
ДНК в составе хромосом чрезвычайно плотно и сложно упакована, и традиционно считалось, что для осуществления процессов матричного синтеза (транскрипция и репликация) эта упаковка мешает и должна быть нарушена в масштабе довольно больших по размеру хроматиновых доменов.
Идентифицировать эти домены и проанализировать их структурную организацию с высоким пространственным разрешением, не нарушив при этом их естественной структуры, было достаточно сложно.
«Мы предложили метод маркировки работающих генов, основываясь на различиях активных и неактивных хромосомных участков во время репликации из ДНК.
Так, активно работающий участок, так называемый эухроматин, реплицируется в самом начале синтетического периода клеточного цикла, а «молчащий» — гетерохроматин — во второй его половине, — комментирует Игорь Киреев.
— Наш метод позволил осуществлять недеструктивные (неразрушающие) исследования хроматина.
Он основан на комбинации мечения новосинтезированной (дочерней) ДНК методами click-химии с последующей детекций продуктов реакции методами корреляционной флуоресцентной микроскопии с суперразрешением и иммуноэлектронной микроскопии. Иными словами, одну и ту же молекулу в данной клетке мы можем видеть и в оптический, и в электронный микроскоп».
Используя данный подход, ученые сделали два неожиданных наблюдения. Во-первых, «работающий» хроматин, вопреки традиционным представлениям, сохраняет весьма высокую степень упаковки, поскольку он представлен высоко структурированными хроматиновыми фибриллами высшего порядка — хромонемами.
Во-вторых, оказалось, что ДНК в составе хромонем обладает высокой структурной пластичностью, то есть способна как бы «перетекать» из одного участка хромонемы в соседний. При этом общая плотная структура хромонемы не изменяется.
Эти наблюдения не укладываются в рамки существующих теорий о пространственной организации хромосом, но в то же время позволяют высказать новые гипотезы о механизмах передачи эпигенетической информации в процессе клеточного деления.
«Мы сделали предположение о том, что в новой дочерней клетке структуры хроматина могут перемещаться внутри нее, а не являться фиксированными, взаимодействуя с той ДНК, которая еще не удваивалась и «помнит все», а также содержит необходимые молекулярные компоненты для восстановления утраченной эпигенетической информации», — продолжает Игорь Киреев.
Другой озвученный в статье вывод состоит в том, что структурная организация генома не является жесткой иерархией. Конечно, есть некие последовательные уровни организации ДНК.
Раньше думали, что должна быть четко зафиксированная система, которая однозначно приводит к формированию хромосом.
Сейчас оказывается, что все может быть по-другому: есть некие принципы построения, но внутри заданных границ ДНК обладает некоторой свободой и пластичностью.
«Дальнейшее развитие исследований состоит, во-первых, в переходе к анализу индивидуальных хромосомных локусов, которые мы планируем маркировать (выделять) на оптическом и электронно-микроскопическом уровнях при помощи технологии TALE, а во-вторых, в разработке еще более нативных методов in vivo мечения, совместимых с такими передовыми технологиями, как криоэлектронная микроскопия», — делится планами на будущее Игорь Киреев.
Ученые надеются вплотную приблизиться к расшифровке принципов пространственной организации ДНК, используя прямые методы анализа при помощи визуализации способов упаковки хроматина с высоким разрешением.
В практическом плане исследования позволят выяснить структурные аспекты эпигенетического контроля генной экспрессии и, возможно, подсказать пути его регулирования, что позволит разрабатывать более эффективные подходы для терапевтических воздействий, например при борьбе с раком и старением — состояниями, в которых очень ярко выражена «эпигенетическая» компонента.